
Download
C h r o m a C * = a * 2 − b * 2 1 Δ E * a b = L 0 * − L * 2 + a 0 * − a * 2 b 0 * − b * 2 3 
ORIGINAL ARTICLE
Integrated taxonomic characterization, postharvest quality evolution, and sustainable conservation of truffle species from Calabria (Southern Italy)
Adriana Chiappetta1, Vincenzo Sicari2*, Rosa Tundis3, Antonio Gattuso2, Roberta Pino3, Michele Pellegrino3, Kevin Garofalo4, Innocenzo Muzzalupo4*, Monica Rosa Loizzo3
1Department of Biology, Ecology and Earth Sciences, University of Calabria (UNICAL), Ponte Bucci, Italy;
2Department of Agraria, “Mediterranea” University of Reggio Calabria (UNIRC), Cittadella Universitaria, Località Feo di Vito, Reggio, Calabria (RC), Italy;
3Department of Pharmacy, Health and Nutritional Sciences, University of Calabria (UNICAL), Rende, Italy;
4Research Center for Forestry and Wood, Council for Agricultural Research and Economics (CREA), Settimio Severo, Rende (CS), Italy
Abstract
Truffles are fungi, usually globular and found underground, typically located using trained dogs. The most -consumed species belong to the genera Tuber, Terfezia, and Tirmania. Southern Italy produces both black and white truffles. Truffles are characterized by a short shelf life due to the loss of quality as a consequence of microbial spoilage and enzymatic browning. This work aimed at the taxonomic identification, by using a molecular approach, of three truffle species, such as Tuber mesentericum Vittad. (TM), Tuber borchii Vittad. (TB), and Tuber aestivum var. uncinatum Chatin (TU), collected from the Calabria region and the evolution of truffle’s quality parameters for 28 days in two storage conditions (under rice and vacuum). The total phenols content (TPC) and total flavonoids content (TFC), degradation kinetics, as well as antioxidant activity were also assessed. Generally, the vacuum packaging proved to be more effective in maintaining the sensory and nutritional quality of TB and TU, whereas the results for TM were more variable, showing greater sensitivity to microbial proliferation under anaerobic conditions. Kinetics results evidenced that the degradation of both TPC and TFC does not depend on the different preservation methods but on the species of truffle object of investigation. Principal Component Analysis does not allow us to establish the advantage of the vacuum preservation method compared to that under rice. Therefore, the use of rice as a matrix to be recovered at the end of the preservation process, edible and rich in truffle aroma, can represent an interesting aspect for the consumer.
Key words: Italian truffles, morphological and molecular characteristics, rice, shelf life, truffles quality, vacuum
*Corresponding Authors: Vincenzo Sicari, Department of Agraria, “Mediterranea” University of Reggio Calabria (UNIRC), Cittadella Universitaria, Località Feo di Vito, Reggio, Calabria (RC), Italy. Email: vincenzo.sicari@unirc.it; Innocenzo Muzzalupo, Research Center for Forestry and Wood, Council for Agricultural Research and Economics (CREA), Settimio Severo, Rende (CS), Italy. Email: innocenzo.muzzalupo@crea.gov.it
Academic Editor: Prof. Angela Zinnai, University of Pisa, Italy
Received: 15 December 2025; Accepted: 20 April 2026; Published: 15 May 2026
© 2026 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Truffles are the fruiting bodies of ascomycete fungi that establish a symbiotic relationship with the roots of -specific trees, such as oak and hazel (Hall et al., 2007; Liu et al., 2017).
This mutualistic association benefits both partners: the tree provides carbohydrates necessary for truffle development, while the fungus enhances the tree’s absorption of water and essential minerals (Lee et al., 2020).
Tubers are usually globular in shape with a wrinkled external surface (Liu et al., 2017). Normally, specially trained dogs are used to locate this type of underground mushroom (Zambonelli et al., 2016). Not all truffle species are edible. Only the fruiting bodies of some (mostly belonging to the genera Tuber, Terfezia, and Tirmania) are edible (Paolocci et al., 2004; Patel et al., 2012). Among them, the most widely consumed are Tuber melanosporum Vittad. , Tuber magnatum Pico , Tuber mesentericum Vittad. (TM), Tuber borchii Vittad. (TB), Tuber brumale Vittad. , Tuber lyonii, and Tuber aestivum Vittad. (Healy et al., 2016).
White and black truffles can be easily differentiated by their morphological characters. The morphological methods traditionally used for truffle classification have recently been improved upon, with the advent of new molecular techniques which provide efficient tools for typing fruitbodies and ectomycorrhiza. Among them, several PCR applications have been particularly successful in identifying mycorrhizal fungi associated with Tuber spp. (Henrion et al., 1994; Lanfranco et al., 1993). In particular, the intragenic internal transcribed spacer (ITS) of the ribosomal genes has been used as the target region. Species-specific internal ITS primers that amplify PCR products of different lengths were selected to distinguish the morphologically similar fruitbodies of fungi for the black truffles (Bonito et al., 2009; Gryndler et al., 2011), or for the white truffles (Amicucci et al., 1998), by aligning their ITS sequences and considering any incidence of intraspecific variability (Leonardi et al., 2020).
In recent years, some morphological and genetic studies have been carried out, aimed at confirming and ascertaining a greater spread in the regional territory not only of more “noble” species but also new species, which can be significantly important from a naturalistic point of view and above all in terms of increasing biodiversity (Iotti et al., 2007; Mello et al., 2006).
Calabria, a region in southern Italy, boasts a rich and varied biodiversity shaped by its varied landscapes, from rugged mountains to pristine coastlines. The region is home to unique ecosystems, including the Pollino, Sila, and Aspromonte National Parks, which host a wide array of flora and fauna. Calabria’s Mediterranean climate supports rare plant species while its forests and wetlands provide habitats for wolves, wildcats, and golden eagles. In Calabria, the presence and spread of truffles have been known for years. Findings of fruitbodies are becoming more frequent, and their macro and microscopic morphologies have given rise, in recent years, to increasing doubts and discussions among truffle harvesters to arrive at the correct taxonomic identification of many samples (Spanò et al., 2024).
Truffles are considered one of the most luxurious and sought-after foods in the world, due to their organoleptic and rare and seasonal characteristics (Reale et al., 2009). However, these products have a notoriously short shelf life due to fast postharvest degradation. The loss of volatile organic compounds, microbial spoilage, and enzymatic browning significantly impact their sensory qualities, making effective storage solutions crucial. Understanding the factors influencing truffle shelf life is essential for improving storage methods and extending their usability without compromising quality.
Temperature, humidity, and oxygen exposure play crucial roles in truffle preservation. Research suggests that refrigeration at 0–4°C extends shelf life up to 10–14 days while minimizing microbial spoilage (Saltarelli et al., 2008). However, prolonged cold storage can alter the biochemical composition, reducing aromatic intensity (Epping et al., 2024). Vacuum packaging and modified atmosphere packaging (MAP) have been explored as alternatives, with studies showing that MAP can retain truffle aroma longer by slowing oxidation and microbial growth (Li et al., 2024; Rivera et al., 2010). However, historically, truffles have been stored using simple techniques such as dry storage (Paper Towels, Cloth Wraps) and immersion in rice. Each method has benefits but also significant limitations. Rice storage, often used in home settings to absorb moisture, accelerates aroma depletion by drawing out VOCs (Strojnik et al., 2020). Nevertheless, interest in this conservative mode has increased due to its sustainability; once rice has completed its task, it can itself be consumed.
The truffle in Calabria is harvested mainly in the province of Cosenza, between the Pollino and the Sila massifs. The right soil, at a specific altitude, and a favorable climate are the characteristics of the hilly woods scattered between the mountains and the Calabrian plain, ideal for black truffle species such as T. aestivum var. uncinatum Chatin (TU) and TM. On the other hand, the prized white truffle (T. magnatum Pico) and the “bianchetto” truffle (TB) prefer moist and shady flat areas, and further toward the sea (Spanò et al., 2024).
This study aimed to provide a comprehensive evaluation of the quality evolution and shelf life of three truffle species (TB, TU, and TM) harvested in the Calabria region (Southern Italy), by integrating taxonomic characterization with an in-depth assessment of postharvest preservation strategies. Specifically, the work pursued a dual objective. First, an integrated morphological, histological, and molecular approach based on ITS rDNA sequencing was applied to confirm the taxonomic identity of the collected ascocarps, contributing to the knowledge of truffle biodiversity in a geographically underexplored area with high ecological value. This aspect addresses the increasing need for accurate species identification, which is essential for both biodiversity conservation and commercial traceability. Second, the study aimed to compare the effectiveness of two preservation methods commonly used in practice, traditional storage under rice and vacuum packaging, on the physicochemical, microbiological, phytochemical, antioxidant, and sensory qualities of fresh truffles during refrigerated storage (4 ± 1°C) over 28 days. To this end, changes in weight loss, firmness, pH, color parameters (CIELAB), microbial load, organic acid profile, total phenolic and flavonoid contents, antioxidant activity, and sensory attributes were systematically monitored. By adopting a multidisciplinary analytical approach, this research sought to elucidate species-specific responses to storage conditions and to identify the main factors influencing quality degradation and consumer acceptability. Ultimately, the work aims to provide scientifically sound and practically applicable information to support the development of more effective and sustainable postharvest handling strategies for fresh truffles, enhancing their commercial value while preserving their distinctive sensory and functional properties.
Materials and Methods
Chemicals and reagents
The chemicals and reagents used for the experimental research were purchased from Sigma-Aldrich S.p.a. (Milan, Italy). Analytical-grade solvents and buffered peptone water (BPW) were obtained from VWR International s.r.l. (Milan, Italy).
Source of fungal material
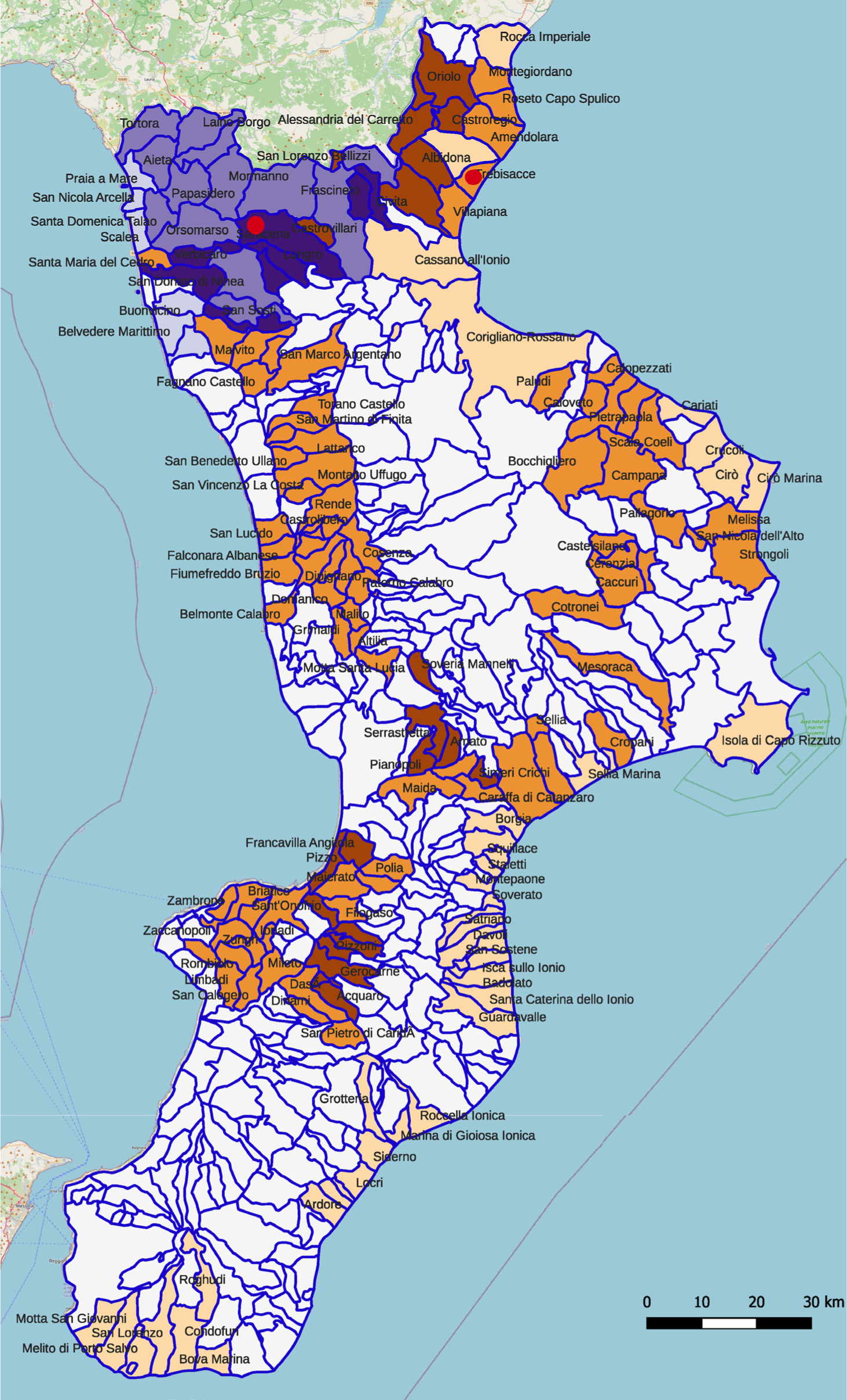
Truffles were collected from two locations in the Calabria region (Pollino area and coast of the upper Ionian Cosenza, Figure 1). The fruitbodies were identified by morphological methods, according to Pegler et al. (1993).
Figure 1. Map of the Calabria Region divided by municipalities, indicating potential truffle-producing areas using different -colors: Beige: possible presence of Tuber borchii; Light brown: possible presence of T. borchii and Tuber aestivum; Dark brown: possible presence of T. aestivum; Light gray: possible presence T. borchii and Tuber uncinatum; Dark gray: possible presence of T. borchii, T. mesentericum, and T. uncinatum; Purple: possible presence T. mesentericum and T. uncinatum. The two truffle sampling points are marked in red: Saracena for T. uncinatum and T. mesentericum, and Trebisacce for T. borchii. Map created by Gilberto Bragato, senior researcher at CREA, considering the soil characteristics and forest species present in the Calabrian territory.
The fruitbodies used in this study belonged to the species TB, TU, and TM.
T. borchii Vittadini 1831 (whitish truffle or Marzuolo truffle, TB). Specimens examined: Italy, Calabria Region, Cosenza Province, Trebisacce County, in soil under Aleppo pine forest (Pinus halepensis), at an altitude of 215 m above sea level, March 4, 2024 (Figure 1).
T. aestivum Vittadini var. uncinatum (Chatin) (hooked truffle, TU). Specimens examined: Italy, Calabrian Region, Cosenza Province, Saracena County, Novacco Plan Location, in soil under beech forest (Fagus sylvatica), at an altitude of 1315 m above sea level, March 4, 2024 (Figure 1).
T. mesentericum Vittadini 1831 (ordinary black truffle or Bagnoli truffle, TM). Specimens examined: Italy, Calabria Region, Cosenza Province, Saracena County, Novacco Plan Location, in soil under beech forest (Fagus sylvatica), at an altitude of 1315 m above sea level, March 4, 2024 (Figure 1).
Histological examination, ascocarps, and ascospores of truffles
For microscopic analysis, truffles were cut using the vibratome (Leica VT1000E, Bensheim, Germany). 100 µm sections were placed on a microscope slide, covered with distilled water (~300 µL per slide) and observed under a stereomicroscope (Optech-Optical technology GZ 808 series equipped with Visiocam 5000 ISHD). Fungal taxa were identified via visual assessment based on spore morphology and appearance (Lacey and West, 2007). Analysis was performed using bright-field illumination microscopy (Leica DRMB, Milan, Italy), and images were taken using the digital camera Leica DFC 320 (Leica, Milan, Italy) (Bruno et al. 2009). Spores (n. 30) were counted and measured for three random fields of view per sample with 40X magnification. Images were analyzed by ToupView Analyze software (Version 4.11), which provided measurement functions and image enhancement options. In the taxonomic descriptions of species, L and W refer to the length and width, respectively, of ascospores ± standard deviation; and “Q” refers to the average of the L/W ratio of all ascospores ± -standard deviation.
DNA isolation, amplification, and sequencing
Genomic DNA was isolated from fresh ascocarps (1.5 mg), as described by Paolocci et al. (2004) with some modifications. The tissue was crushed in -liquid nitrogen in a 2 mL microcentrifuge tube, using a sterilized micropestle. The ground tissue was suspended in 150 µL buffer containing 200 mM Tris-HCl, pH 7.5, 250 mM NaCl, 25 mM EDTA, and 0.5% SDS, vortexed for 10 s and kept at room temperature until all samples were extracted. The extracts were centrifuged for 10 min at 10,000 × g, and the supernatants were transferred to a new microcentrifuge tube and precipitated in an equal volume of isopropanol for 20 min at −20°C. The DNA was pelleted by centrifugation for 15 min, vacuum-dried, and resuspended in 50 µL double-distilled nuclease-free water. Resuspension of the pellet was occasionally facilitated by warming at 65°C for 5 min. When clumps were present, a further 5-min centrifugation was performed, and the supernatant was carefully transferred to a new tube. The DNA concentration in final extracts was measured at 260 nm using a NanoDrop spectrophotometer (Thermo Scientific, Wilmington, DE). Undiluted extracts were used directly as a template in PCR.
DNA of the Internal Transcribed Spacer (ITS) regions were amplified using the universal primers ITS1: 5’-TCC GTA GGT GAA CCT GCG G-3’ and ITS4: 5’-TCC TCC GCT TAT TGA TAT GC-3’ (White et al., 1990). PCRs were prepared as follows: a reaction tube contained 50 ng target DNA, 2.5 µL reaction buffer, 2.5 µL 10 mg/mL BSA, 0.5 µL 100 µM forward primer, 0.5 µL 100 µM reverse primer, 2.5 mL 1 mM dNTPs, 1.5 µL 25mM MgCl2, and 0.2 µL (5 U/µL) of Taq DNA polymerase. The thermocycle was run 3 min at 94°C, and then 30 cycles as follows: 30 s at 94°C, a gradient of annealing temperature from 55 to 70°C for 30 s and 2 min at 72°C. An elongation period of 10 min was allowed at 72°C before cooling or removing the tubes. PCR products were analyzed by 2100 Bioanalyzer (Agilent Technologies, Waldbronn, Germany) with the DNA 1000 kit, which gave an estimate of the amplified product length (Chiappetta et al., 2017). PCR amplification from single ascomata was -carried out at least in triplicate.
All amplified DNA samples were assayed on a Qubit fluorimeter (Invitrogen Thermo Fisher) and diluted with distilled water to a concentration of 5 ng/µL and 20 µL (100 ng total DNA). All samples were placed on 96-well PCR microplates and sent to Genomix4Life (Salerno, IT) for sequencing. Sequencing reactions were performed on purified PCR products obtained with ITS1 and ITS4 primers. Sequences of the ITS regions, including the 5.8S gene of rDNA, were compared to those of the same species present in GenBank (accession numbers: KT215193 (TB) (Baral et al., 2018), AF516786 (T. aestivum), and AF516798 (TM) (Amicucci et al., 1998; Paolocci et al., 2004). BLAST searches (blastn: https://blast.ncbi.nlm.nih.gov; Altschul et al., 1997) were conducted for the partial ITS of nuclear rDNA sequences against the International Nucleotide Sequence Database (GenBank: http://www.ncbi.nlm.nih.gov/).
Sample preparation and experimental design
Two different preservation methods were compared: the traditional one, which involves placing truffles inside glass containers under rice, and one based on vacuum preservation. The two methods were applied to the three truffle species (TM, TB, and TU) collected in the Calabria region (Italy). Truffles were cleaned with a soft and wet brush, rinsed with running water and dried under a laminar flow hood. Carpophores with a soft consistency, parasitized or severely damaged during collection (by tools or dogs during the search) were discarded. After selection, truffles were stored at 4°C for 24 h before packaging. The samples were divided into two groups:
-
Under rice (UR) – Rice-packed using “Originario” rice (Despar, Italy);
-
Under vacuum (UV) – Vacuum-packed, using multilayer food-grade polyethylene bags (350 ×180 ×150 mm) (OLPACK 510, Interprise Brussels S.A., Belgium).
Both groups were stored at 4 ± 1°C for 28 days. Each treatment was replicated three times (each replicate consisted of one package containing 5 truffles), and the samples were analyzed at Day 0 (t0) and at regular intervals of 7 days until Day 28 (t28). To ensure the statistical reproducibility of the samples for each species, we collected approximately 150 samples per species.
Quality and CIELAB parameters
Weight loss during storage of packaged truffles was determined every 7 days using a PCE-BSK 310 laboratory balance digital scale (PCE-BSK 310, PCE Instruments, Capannori (LU), Italy).
Truffles were removed from the trays, and weight loss was immediately determined (AOAC-Official Methods of Analysis 2000). Results were expressed as WL (%) = (IW – FW)/IW × 100, where (IW): initial weight, (FW): final weight.
The pH was measured using a pH meter with a glass electrode (Crison 2001, Crison Instruments, Barcelona, Spain) inserted into homogenized truffles, 10 g + 90 mL of sterile physiological water. The measurements were carried out in triplicate for each sample and expressed as mean ± standard deviation. Changes in truffle firmness were investigated using a texture analyzer (mod. TA.TX2, Stable Microsystems, Surrey, UK).
The Texture expert program version 1.22 was used for data analysis. A 4 mm diameter cylinder probe (mod. P/4) was used for the puncturing test. Firmness was defined as the force recorded in a force–time curve obtained from the texture analyzer at a depth of penetration of 5 mm during the compression of the truffle by the cylinder probe. The results of the penetration test were expressed from the time versus force curves in N/mm. Firmness was determined as the maximum force.
CIELab parameters (a*, b* and L*) were measured at 25°C using the PCE CSM-4 colorimeter (PCE, Lucca, Italy). Data were expressed as a higher saturation of color or Chroma (C*) (1). Hue angle (h*) (2), the qualitative indicator of color, was also calculated. A higher hue angle represents a lesser yellow character in the assays. An angle of 0° or 360° represents red Hue, while angles of 90°, 180°, and 270° represent yellow, green, and blue hues, respectively (Sicari et al., 2024).
Hue angle (h*)= tan-1 (b*/a*)(2)
The overall change of truffle color was expressed as ∆E*ab parameter, calculated following the equation:
where the index “0” indicates the sample at time 0, whereas the letters without an index correspond to the parameters of the samples during storage.
Microbiological analysis
Microbial analyses of truffles, which included the enumeration of mesophilic aerobic microorganisms, were determined before and after the application of the vacuum and/or rice treatments and weekly, until the end of the shelf life of the truffle batches (0, 7, 14, 21, 28 days).
Each sample was decimal diluted in 0.1% BPW (sterile distilled peptone water: Merck, Germany) and homogenized using a laboratory Smasher Stomacher (AES, Laboratore) for 2 min at 250 rpm. The BPW was used for subsequent serial dilutions. One milliliter of each dilution was pour-plated on plate count agar (PCA, Sigma Aldrich, Italy) for mesophilic aerobic plate counts and incubated for 48 h at 30°C. The data are presented as the mean of three samples and expressed as log cfu/g.
Evaluation of organic acids during storage
The content of organic acids (citric, fumaric, malic, oxalic, succinic, and tartaric acids) was determined following the method previously reported by Sicari et al. (2024), with some modifications. The homogenized truffles were diluted to 10-fold with ultrapure water, and after centrifugation at 4000 × g for 15 min, the supernatants were separated and filtered through a 0.45 µm PTFE syringe filter (Supelco, Milan, Italy). The quantification of specific compounds was carried out by a Knauer chromatograph equipped with a UV detector operated at 210 nm and coupled with an Acclaim OA5 column (4 mm i.d. × 250 mm length × 5 µm particle size). A mobile phase of 0.2 M KH2PO4 was used for the separation with a flow rate of 0.5 mL/min. An external standard calibration curve was used for the identification and quantification of components. Five injections were made for each calibration level. The r2 values were between 0.998 and 0.999 for linear regression of the curves of external calibration standards. Data -processing was carried out with the support of Clarity Chromatography Software 8.1 (GmbH, Tübingen, Germany), and results were expressed as mg of acid/100 g of sample.
Ultrasound-assisted extraction procedure
Truffles packaged in different packaging systems (250 g) were subjected to an ultrasound-assisted maceration process using ethanol/water (9:1 v/v, 500 mL × three times) as solvent in a Branson model 3800-CPXH water bath (Branson, Milan, Italy) with a frequency of 40 kHz at 25°C for 50 min. The extraction procedure was repeated thrice, and after each extraction cycle, the solutions were filtered, and the solvent was evaporated. The obtained extracts were used for phytochemical content determination as well as biological activity assessment.
Total phenols and flavonoids content
The Folin–Ciocalteu method was applied to estimate the total phenol content (TPC) as previously reported (Tenuta et al., 2020). Samples at a concentration of 1.5 mg/mL (0.1 mL) were mixed with a solution of Folin–Ciocalteu reagent (0.5 mL) and water (1 mL), then 1.5 mL of 20% sodium carbonate was added, and the mixture was incubated at room temperature. The absorbance was measured at 765 nm using a UV-Vis (Jenway 6003, Carlo Erba, Milan, Italy). The TPC was expressed as mg of chlorogenic acid equivalents (CAE)/g fresh weight (FW).
For the total flavonoids content (TFC) determination, a method based on the formation of a flavonoid–aluminum complex (Tenuta et al., 2020) was used. Truffle extract was mixed with aluminum chloride solution (2%) in a 1:1 ratio and incubated at room temperature for 15 min.
The absorbance was measured at 510 nm, and the TFC was expressed as mg quercetin equivalents (QE)/g FW.
Degradation kinetics of TPC and TFC
The impact of storage conditions on TPC and TFC was investigated using a mathematical kinetic model (Sicari et al., 2024). The first-order degradation kinetic was applied following the equations:
–In (Ct/C0) = kt
t1/2 = In (2)/k
where C0 is the TPC and TFC at Day 0, and Ct is the concentration of both phenols and flavonoids after 7, 14, 21, and 28 days of storage.
Antioxidant activity
The antioxidant activity of truffles during storage was assessed by ABTS, DPPH, β-carotene bleaching test, and FRAP test.
The ABTS radical scavenging assay was performed following the procedure reported by Loizzo et al. (2021). The ABTS solution (7 mM) was mixed with potassium persulfate (2.45 mM) to obtain an ABTS+ radical cation solution. After 12 h, this latter solution was diluted with ethanol to a final absorbance of 0.70 at 734 nm using a Jenway 6003 UV-Vis spectrophotometer. Subsequently, 2 mL of diluted ABTS+• solution was added to the extract (25 µL) at concentrations from 400 to 1 µg/mL. After 6 min, the absorbance was read at 734 nm. The ABTS removal capacity was calculated as follows: ABTS removal activity (%) = ([A−A0]/A0) 100, where A0 is the absorbance of the control reaction, and A is the absorbance in the presence of the extract.
To perform the DPPH radical scavenging assay, a previously published procedure (Loizzo et al., 2021) was used. Truffle’s extract at different concentrations (1–1000 µg/mL) was added to the DPPH solution, and after 30 min, the absorbance was measured at 517 nm. The DPPH radical scavenging activity was calculated following the equation: DPPH radical scavenging activity (%) = (1 [absorbance of sample with DPPH absorbance of sample without DPPH]) × 100. Ascorbic acid was used as a positive control in both radical scavenging assays (ABTS and DPPH).
The β-carotene bleaching test was used to investigate the potential of truffles to inhibit peroxidation following the procedure previously described (Loizzo et al., 2021). Briefly, Tween 20, linoleic acid, and β-carotene were mixed. After evaporation of the solvent, the emulsion was assigned in the 96-well microplate containing truffle’s extract at different concentrations (2.5–100 µg/mL). After incubation at 45°C for 30 min, absorbance was measured at 470 nm. Propyl gallate was used as the positive control.
The FRAP test is based on the redox reaction that involves the TPTZ (2,4,6-tripyridyl-striazine)-Fe3+ complex. FRAP reagent was mixed with truffle extract dissolved in ethanol at a concentration of 2.5 mg/mL. After 30 min of incubation at 25°C, the absorbance was read at λ = 595 nm using a UV-Vis Jenway 6003 spectrophotometer (Carlo Erba, Milan, Italy). The result was expressed as FRAP value in μMFe (II)/g. BHT was used as a positive control (Loizzo et al., 2021).
Sensory analysis
Truffles were removed from the trays and left at room temperature for 1 h to allow the product to reach tasting temperature. A panel of six trained judges, recruited from the CibusLab Association (CibusLAB – Sensory analysis laboratory for the protection and enhancement of Italian agrifoods, Cosenza, Italy), evaluated truffle quality in terms of texture, aroma, and flavor using a 9-point structured scale (Rivera et al., 2010).
Texture was evaluated by gently placing the truffles between the ring and index fingers and applying slight pressure. The following scores were used for texture: hard = 9, moderately hard = 7, moderately soft = 5, soft = 3, and unacceptable = 0. Aroma was assessed using the following scale: fully typical aroma = 9, moderately full aroma = 7, moderate aroma = 5, slight aroma = 3, and no typical truffle aroma = 0. For flavor evaluation, truffles were sliced (approximately 1 mm), and judges assessed the samples using a scale where 9 = fully typical flavor, 7 = moderately full flavor, 5 = moderate flavor, 3 = slight flavor, and 1 = no typical flavor.
Truffles were considered unmarketable when aroma, flavor, and texture scores reached values close to or lower than 5. Sensory data were collected and processed using the Smart Sensory Box software (Smart Sensory Solutions S.r.l., Sassari, Italy). The panel leader collected the evaluation sheets, entered the data into the software, and performed the statistical analysis based on the calculation of median values and the robust coefficient of variation (CVr%). Only sensory results showing CVr ≤ 20% were considered acceptable, in accordance with ISO guidelines. In cases of anomalous responses, judges were asked to revise their evaluation sheets and, when necessary, to repeat the sensory test.
Statistical analysis
Data processing, calculation of mean values, standard deviation (SD), relative standard deviation, linear regression, and Pearson’s correlation coefficient (r) were initially performed using Microsoft Excel 2010 (Microsoft Corporation, Redmond, WA, USA). Results were expressed as mean ± SD (n = 3), unless otherwise stated.
Statistical analysis was carried out using SPSS software for Windows, version 22.0 (SPSS Inc., Chicago, IL, USA) using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test, and multivariate analysis to evaluate the effects of storage time and preservation technique, as well as their interaction, on all quantitative parameters (Table 10). When significant effects were detected, mean comparisons were performed using Tukey’s post hoc test. Differences were considered statistically significant at P < 0.05 and highly significant at P < 0.01.
The inhibitory concentration at 50% (IC50) was calculated using GraphPad Prism version 4.0 for Windows (GraphPad Software, San Diego, CA, USA).
Sensory analysis was conducted by a trained panel of six judges, and each sample was evaluated in triplicate at each storage time. Sensory data were expressed as median values, and data variability was assessed using the robust coefficient of variation (CVr%) calculated by Smart Sensory Box software. Only sensory attributes with CVr ≤ 20% were considered reliable and included in the analysis. Given the ordinal nature of sensory scores and the limited panel size, robust nonparametric statistics were applied in accordance with standard sensory analysis guidelines.
Principal Component Analysis (PCA) was performed using SPSS software for Windows, version 22.0 (SPSS Inc., Chicago, IL, USA) on the entire dataset merged into a single multivariate model.
Results and Discussion
Morphological characterization of tuber species
The integrative taxonomic approach, based on macroscopic and microscopic traits, allowed the identification of three species occurring in the Calabria Region: TB, TU, and TM (Figures 1–3).
Figure 2. Morphological characteristics of external (A, B, C) and internal A’, B’, C’ surfaces of truffle fruitbodies, (A) Tuber -borchii; (B) Tuber aestivum var. uncinatum; (C) Tuber mesentericum (space bar = 1 cm).
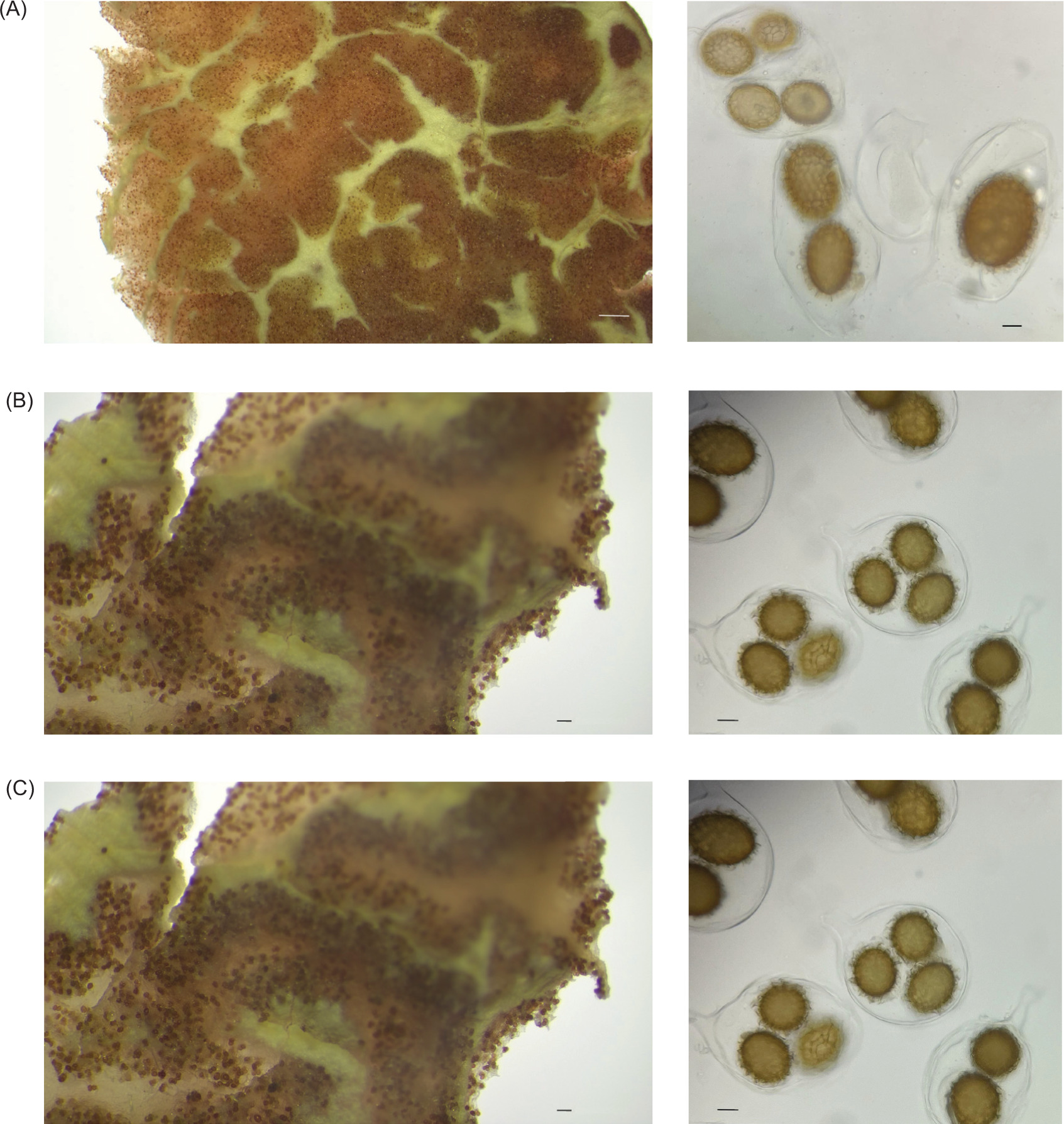
Figure 3. (A, B, C) Stereomicroscope and (A’, B’, C’) optical microscope images of truffle fruiting bodies. (A) Tuber borchii gleba (scale bar = 1 mm); (A’): T. borchii asci and ascospores (scale bar = 10 µm); (L= 28.9 ± 2,4; W= 38.6 ± 5.3; Qm = 0.75 ± 0.09). (B) Tuber aestivum var. uncinatum gleba (scale bar = 100 µm); (B’): T. aestivum var. uncinatum asci and ascospores (scale bar = 10 µm); (L = 28.1 ± 1.4; W = 20.1 ± 1.4; Qm = 1.39 ± 0.14). (C) Tuber mesentericum Vitt. gleba (scale bar = 100 µm); (C’): T. -mesentericum Vitt. asci and ascospores (scale bar = 10 µm); (L = 40.5 ± 8.2; W = 30.2 ± 4.2; Qm = 1.34 ± 0.29).
Tuber borchii Vittadini 1831 (whitish truffle or Marzuolo truffle, TB)
Ascomata: Mostly tuberiform, rounded, and lobed-gibbous. It ranges in size from a few millimeters to 3–4 cm in diameter, with a maximum size of 7 cm, and weighs from a few grams to over 150 g. Peridium: Smooth and variable in color from whitish or chalky white to light ochre and even reddish brown, with frequent rust-red spots. In young specimens, the surface is pubescent due to the abundant presence of short, rigid hyphae 3–5 µm in size. In adult fruit bodies, the hairs are generally less abundant; the peridium appears smoother and has a pseudoparenchymatous structure with irregular, oblong cells. Gleba: In immature fruiting bodies, it is firm and whitish; in mature ones, it turns ochre, pink to reddish brown, sometimes with violet shades. The sterile veins are sometimes broad with an undefined outline, while in other cases, they are thin, branched, tortuous, and very distinct (Figure 2A). Asci: They are globose, ellipsoidal, or subclavate, lack a stalk, and contain 1–4 spores. Ascospores: They are yellow–brown, ellipsoidal, subglobose, and isopolar, with very broad, rounded poles. They measure 24–33 (L) × 30–45 (W) µm; Q(L/W) = 0.73–0.80. In monosporic asci, the spore is very large. (Figure 3A). Episporium: It consists of very small, generally hexagonal or pentagonal, mostly regular alveoli, which give the episporium a reticulated appearance. The width of the mesh varies and is independent of the size of the spores: the smallest ones measure about 4 µm in width, and the largest ones can reach 7 µm in diameter. Aroma: It is strong, slightly pungent, garlicky, and pleasant in small amounts.
Tuber aestivum Vittadini var. uncinatum (hooked truffle, TU)
Tuber aestivum var. uncinatum is a prized species of truffle in the Tuberaceae family. From a morphological point of view, it has the following characteristics: Ascomata: The fruiting body of this truffle is generally globose or ovoid but can also have an irregular shape. Its size varies, typically reaching a diameter of 4–10 cm, so the weight can fluctuate from a few grams to 400–500 g. The surface is rough and irregular, with prominent and irregular warts, giving it a rather coarse and knobby appearance. The warts vary in size and contribute to the truffle’s distinct texture. Peridium: The outer layer varies in color from dark grey to black, brown, with some areas exhibiting a brownish tint. At certain maturation stages, it may appear lighter. Its texture is relatively thin compared to other truffle varieties. Warts are generally small or medium, 2–5 mm wide, with triangular lateral faces. Gleba: The inner flesh is compact and light colored, ranging from white to pale yellow, with irregular dark brown branching veins when ripe. These veins are quite noticeable and give the flesh a distinctive appearance (Figure 2B). Asci: Globose with a short peduncle, measuring about 80–100 (L) × 55–75 (W) µm; Q(L/W) = 1.33–1.45), and containing from 1 to 6 spores (Figure 3B). Ascospores: Ellipsoidal or subspherical, irregular, dark brown in color, measuring about 25–30 (L) × 18–22 (W) µm; Q(L/W) = 1.36–1.39 (Figure 3B). Episporium: has large, regular hexagonal and pentagonal alveoli; 9-11 µm wide with very high walls of 4–8 µm. The walls of the alveoli are straight and sometimes frayed. Additional ridges may be present within the alveoli. Aroma: Delicate, hazelnut-like, with earthy undertones.
Tuber mesentericum Vittadini 1831 (ordinary black truffle or Bagnoli truffle, TM)
Ascomata: It is rounded, sometimes with irregular lobes, and almost always featuring a deep depression, giving the fruiting body a kidney-shaped appearance in section. It has a diameter of 2–6 cm, rarely larger, and the weight varies from a few grams to about 100 g. Peridium: It is blackish or dark brown with small warts measuring 2–4 mm in width and 2–3 mm in length, with sharp edges. Gleba: Initially white, at full maturity, it turns gray-brown or brown in various shades (Figure 2C). Asci: They are spherical, with a short stalk, and contain 2–6 spores. Ascospores: They are ellipsoidal in shape, with broad ridges and a brown-translucent color. They measure 27–53 (L) × 23–37 (W) µm; Q(L/W) = 1.43–1.17 (Figure 3C). The episporium is alveolate with medium-sized to small alveoli, rarely completely closed by lateral walls of irregular height that often take on the appearance of jagged ridges. The aroma is very intense but decreases when exposed to air, resembling tar and iodine tincture.
Molecular identification
PCR amplification of ITS regions with primers ITS1 and ITS4 yielded clear products of 654 bp (TB), 689 bp (TM), and 705 bp (TU). BLAST analysis confirmed high sequence similarity with GenBank accessions: 98.63% (TB), 98.83% (TM), 98.86% (TU), supporting morphological determinations (Amadoro et al., 2013; Rivera et al., 2010).
Physicochemical and parameters
In recent years, demand for truffles has steadily increased at both the European and global levels. However, this growing demand poses significant challenges for approval and conservation, given the seasonal nature of truffle production and their stable harvest periods. Moreover, truffles are highly perishable products (Mannozzi et al., 1994). Their quality and shelf life are influenced by several factors, including pedoclimatic, microbiological, and mechanical conditions. Consequently, preservation strategies must focus on harvesting practices, postharvest handling, and appropriate storage methods.
Traditionally, truffles are preserved using methods such as storage in jars containing brine, brandy, or rice. More recently, the expanding market for fresh truffles has driven research toward innovative technological approaches to extend shelf life. Modified atmosphere packaging (MAP) has been investigated, although it has only partially addressed preservation issues (Rosnes et al., 2003). For instance, TU truffles packaged under MAP with 60% CO2 were successfully stored for up to 3 weeks without significant losses in weight, aroma, or consistency (Massantini et al., 2009). Rivera et al. (2010) evaluated the storage of fresh TM and TU truffles in microperforated polypropylene bags under atmospheric gas mixtures at different temperatures (4, 10, and 23°C). The best results were obtained at 4°C using microperforated films with two holes (90 × 50 μm), which maintained an internal atmosphere of 15% CO2 and 7% O2. Under these conditions, truffles showed reduced microbial counts, better weight retention, and high sensory scores, with shelf life extensions of 28 days for TM and 21 days for TU.
In the present study, we have investigated the effects of two traditional preservation methods: storage under rice and vacuum packaging on Calabrian truffles (TB, TM, and TU). Samples were stored either under rice (“Originario,” Despar, Italy) or vacuum-packed in food-grade multilayer polyethylene bags (OLPACK 510, Interprise Breussel S.A., Belgium) and kept at 4 ± 1°C for 28 days. Each treatment was performed in triplicate, with analyses conducted at Day 0 and at 7-day intervals thereafter.
Truffles are characterized by a high moisture content (approximately 74%), which strongly influences freshness, flavor, and storage performance. Excess internal humidity promotes deterioration, leading to reduced sensory quality, rapid weight loss, and visible shrivelling. Weight loss ranged from −5.86 to −6.23% for TUUR and TBUR, and from −6.01 to −6.56% for TUUV and TBUV, respectively (Table 1).
Table 1. Truffles weight (g) during storage.
| Weight | t0 | t7 | t14 | t21 | t28 | Sign. |
|---|---|---|---|---|---|---|
| TUUR | 41.20 ± 1.21a | 40.95 ± 1.32a | 40.20 ± 2.04a | 39.80 ± 0.89a | 28.19 ± 0.77b | ** |
| TUUV | 42.77 ± 1.11a | 41.20 ± 1.43b | 40.20 ± 2.01bc | 40.05 ± 2.65bc | 38.34 ± 1.09d | ** |
| TMUR | 44.63 ± 2.21a | 44.05 ± 2.02a | 43.05 ± 2.12b | 41.72 ± 1.39c | 38.31 ± 1.65d | ** |
| TMUV | 44.63 ± 2.34a | 42.36 ± 2.09b | 42.36 ± 1.79b | 42.86 ± 1.94b | 42.62 ± 1.73b | ** |
| TBUR | 43.18 ± 2.52a | 40.17 ± 1.66b | 40.12 ± 1.56b | 40.49 ± 1.68b | 40.41 ± 1.57b | ** |
| TBUV | 43.18 ± 1.89a | 41.26 ± 1.78b | 41.86 ± 1.63b | 40.11 ± 2.87b | 40.87 ± 1.49b | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TMUR: Tuber mesentericum under rice; TBUR: Tuber borchii under rice; TUUV: Tuber aestivum var. uncinatum; TMUV: Tuber mesentericum under vacuum; TBUV: Tuber borchii under vacuum. Data are expressed as means ± SD (n = 3). Differences between samples were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant. **P ≤ 0.01.
These results are comparable to those reported by Savini et al. (2020), who observed a gradual increase in weight loss during storage of black truffles at 4°C, regardless of MAP conditions. In that study, weight losses ranged from −6.7 to −9.9% under 40% CO2/60% N2 and air atmospheres, respectively. Other studies have reported lower weight losses following alternative treatments. Daei et al. (2023) demonstrated that a 5% vinegar treatment at 63°C effectively reduced weight loss in Terfezia clavery during storage, while Amadoro et al. (2013) found weight losses of −2.62 and −3.43% after 28 days in control samples and truffles treated with combined decontamination and storage methods, respectively.
Initial pH values of 6.67, 6.64, and 6.69 were recorded for TU, TM, and TB, respectively. As shown in Table S1, a slight decrease in pH occurred during storage, regardless of the preservation method. After 28 days at 4°C, the lowest pH value was observed in TUUR samples (5.86), followed by TMUR (6.01). This reduction is likely associated with changes in microbiological parameters, particularly total bacterial counts (TBCs). These findings are consistent with Amadoro et al. (2013), who reported an initial pH of 6.45 in fresh TU, although a slight pH increase was observed in samples treated with ultrasound or chitosan after storage.
Firmness, a key indicator of truffle quality, was also monitored. High-quality truffles are characterized by high firmness, whereas soft or mushy textures indicate overmaturation. Initial firmness values were 20.33 N for TU, 20.88 N for TM, and 21.67 N for TB. Both storage methods caused a gradual decrease in firmness over time (Table 2). After 28 days, firmness loss ranged from 18.01 to 30.64% in vacuum-packaged truffles (TMUV and TBUV), while higher reductions (25.53–34.98%) were observed in rice-stored samples (TMUR and TBUR). These results confirm firmness loss as a reliable indicator of spoilage and are in agreement with findings reported by Tejedor-Calvo et al. (2024) and Savini et al. (2020).
Table 2. Evolution of firmness parameter (N) in truffles stored in two different conditions.
| Firmness (N) | t0 | t7 | t14 | t21 | t28 | Sign. |
|---|---|---|---|---|---|---|
| TUUR | 20.33 ± 0.45a | 17.79 ± 0.66b | 16.85 ± 0.88c | 15.82 ± 1.01d | 14.02 ± 0.55e | ** |
| TUUV | 20.33 ± 0.87a | 19.02 ± 0.45b | 18.45 ± 0.65c | 17.89 ± 0.99d | 16.02 ± 0.67e | ** |
| TMUR | 21.78 ± 1.03a | 20.24 ± 0.78b | 18.12 ± 0.53c | 17.11 ± 0.79d | 16.22 ± 0.98e | ** |
| TMUV | 20.88 ± 0.89a | 20.06 ± 0.89b | 19.05 ± 0.78c | 18.05 ± 0.93d | 17.12 ± 0.97e | ** |
| TBUR | 21.67 ± 0.56a | 19.44 ± 0.86b | 17.44 ± 0.87c | 15.84 ± 0.82d | 14.09 ± 1.02e | ** |
| TBUV | 21.67 ± 1.64a | 20.12 ± 1.08b | 18.78 ± 0.59c | 16.89 ± 0.74d | 15.03 ± 0.92e | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TMUR: Tuber mesentericum under rice; TBUR: Tuber borchii under rice, and TUUV: Tuber aestivum var. uncinatum under vacuum; TMUV: Tuber mesentericum under vacuum; TBUV: Tuber borchii under vacuum. Data are expressed as means ± SD (n = 3). Differences between samples were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant. *P ≤ 0.05; ** p ≤ 0.01.
Effect of storage on truffles CIELab parameters
The effect of the different storage methods on ascomycete color was evaluated by monitoring CIELab parameters over 28 days of storage. As reported in Table 3, significant differences were observed between vacuum packaging and the traditional storage method for rice.
Table 3. Evolution of chroma (C*) and lightness (L*) in truffles stored in two different conditions.
| t0 | t7 | t14 | t21 | t28 | Sign. | |
|---|---|---|---|---|---|---|
| C* | ||||||
| TUUR | 6.71 ± 0.09e | 7.01 ± 0.09d | 7.52 ± 0.46c | 7.63 ± 0.45b | 7.76 ± 0.78a | ** |
| TUUV | 6.44 ± 0.11e | 6.88 ± 0.22d | 7.41 ± 0.67c | 8.76 ± 0.59b | 10.96 ± 0.66a | ** |
| TMUR | 7.04 ± 0.21e | 8.03 ± 0.67d | 8.90 ± 0.95c | 9.09 ± 0.83bc | 9.72 ± 0.99a | ** |
| TMUV | 7.04 ± 0.43e | 8.40 ± 0.65d | 9.25 ± 0.88c | 13.82 ± 0.72b | 16.00 ± 1.03a | ** |
| TBUR | 8.34 ± 0.23d | 9.30 ± 0.78c | 9.78 ± 0.77c | 10.30 ± 1.06ab | 10.54 ± 1.06a | ** |
| TBUV | 10.13 ± 0.45e | 11.45 ± 0.88d | 12.28 ± 0.78c | 15.46 ± 1.07b | 18.47 ± 1.11a | ** |
| L* | ||||||
| TUUR | 20.32 ± 1.33b | 22.52 ± 1.74a | 17.95 ± 1.46c | 17.57 ± 1.34c | 17.60 ± 1.67c | ** |
| TUUV | 20.32 ± 1.73a | 19.2 ± 1.47ab | 19.02 ± 1.48b | 18.66 ± 1.73b | 17.49 ± 1.89c | ** |
| TMUR | 21.06 ± 1.73c | 25.69 ± 1.45a | 18.92 ± 1.44d | 20.32 ± 1.63c | 22.10 ± 1.78b | ** |
| TMUV | 21.06 ± 1.93a | 20.79 ± 1.78a | 20.24 ± 1.78ab | 17.45 ± 1.89c | 17.01 ± 1.65c | ** |
| TBUR | 23.68 ± 1.03a | 19.57 ± 1.89b | 17.33 ± 1.93c | 18.14 ± 1.98c | 19.76 ± 1.565b | ** |
| TBUV | 25.03 ± 2.03a | 23.22 ± 1.87ab | 23.01 ± 1.73b | 22.73 ± 1.76b | 22.06 ± 1.63b | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TMUR: Tuber mesentericum under rice; TBUR: Tuber borchii under rice, and TUUV: Tuber aestivum var. uncinatum under vacuum; TMUV: Tuber mesentericum under vacuum; TBUV: Tuber borchii under vacuum. Data are expressed as means ± SD (n = 3). Differences between samples were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant. *P ≤ 0.05; **P ≤ 0.01.
Overall, chroma (C*) values increased during storage, regardless of the preservation method applied (Table 3). At Day 0, C* values were 6.71, 7.04, and 8.34 for TU, TM, and TB, respectively. After 28 days, a marked increase in C* was observed, particularly in TM stored under rice (+37.48%). Even greater increases were recorded in vacuum-packaged samples, with C* variations ranging from +51.51% in TUUV to +82.00% in TBUV.
Regarding lightness (L*), truffles stored under rice exhibited a significant reduction in brightness, with decreases ranging from −22.07% in TU to −26.35% in TM. In contrast, vacuum storage resulted in a less pronounced decrease in L*, with reductions between −11.87% in TB and −19.23% in TM (Table 3). The reduction in L* can be attributed to enzymatic browning processes, mainly related to the activation of phenylalanine ammonia lyase and peroxidase following tissue damage during postharvest handling (Shen et al., 2006).
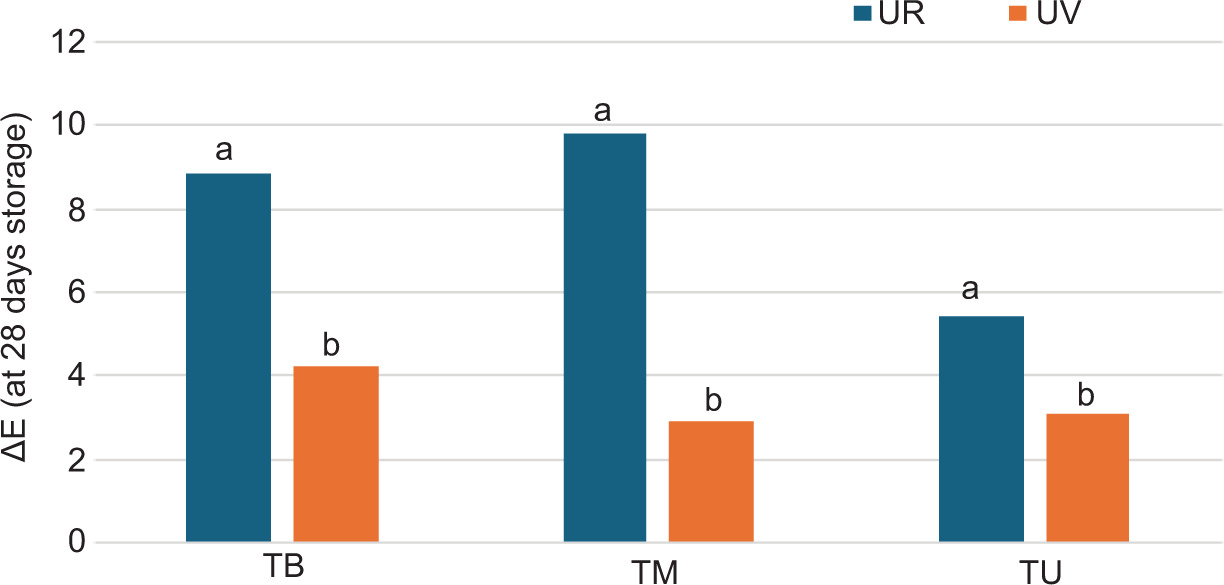
Given that browning is a common phenomenon during food storage, treatment effectiveness was further assessed by calculating the total color difference (ΔE). After 28 days of storage, the samples showed perceptible color changes in both conditions. However, storage in UR resulted in a significant increase in ΔE compared to UV, indicating lower color stability. Sample TM was the most sensitive, while TU showed the best color stability, particularly under UV conditions (Figure 4).
Figure 4. ΔE parameter at 28 days storage at 4°C calculated for truffles storage under rice and under vacuum. TU: Tuber aestivum var. uncinatum; TM: Tuber mesentericum; TB: Tuber borchii. Data are expressed as means ± SD (n = 3). Differences between samples were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant. **P ≤ 0.01.
Microbiological analysis
Members of the Pseudomonas genus, Enterobacteriaceae, lactic acid bacteria, molds, and yeasts are known to be responsible for postharvest truffle spoilage (Rivera et al., 2010). As expected, at the beginning of the experiment (time 0), no significant differences in total microbial count (TBC) were observed among the three truffle species analyzed (TB, TU, and TM). Conversely, in all three species, TBC reached its maximum after 14 days of storage, irrespective of the storage method (vacuum packaging or storage in rice). Notably, after 7 days of storage, TBC was consistently higher in samples stored in rice across all three truffle species (Table S2). At Day 28 (end of shelf life), vacuum-packaged TB samples exhibited a lower microbial load than those stored in rice (1.2 × 108 CFU/g vs 2.7 × 109 CFU/g (Table S2). In contrast, at Day 21, TM and TU showed distinct responses to packaging type: TM samples stored under vacuum exhibited a higher microbial load than those stored in rice (2.49 × 109 CFU/g vs 2.40 × 106 CFU/g), whereas TU samples showed higher microbial counts in rice storage compared to vacuum packaging (2.00 × 109 CFU/g vs 1.92 × 108 CFU/g).
At the end of the storage period (28 days), TU and TM displayed similar microbial growth trends, which were opposite to those observed in TB samples. Unexpectedly, in TU and TM, vacuum packaging resulted in higher TBC than rice storage (TU: 1.42 × 109 CFU/g vs 1.94 × 107 CFU/g; TM: 1.53 × 109 CFU/g vs 1.71 × 107 CFU/g), whereas in TB the opposite trend was observed (1.23 × 108 CFU/g vs 2.71 × 109 CFU/g). This behavior may be attributed to the opportunistic growth of anaerobic microorganisms under vacuum conditions compared to aerobic strains.
Comparable microbial loads have been reported in the literature. Phong et al. (2022) reported total microbial counts of 7.94 and 8.33 log CFU/g in whole and sliced Spanish T. melanosporum, respectively, in agreement with values observed in T. melanosporum harvested in the Marche region (Italy) (Saltarelli et al., 2008). Rivera et al. (2010) also reported high microbial loads in TM and TU, reaching 9.3 and 8.0 log CFU/g, respectively (Figure 5). Several studies have identified Pseudomonas spp. as the dominant microbial group in truffles (Rivera et al., 2010; Sbrana et al., 2002), likely reflecting species-specific microbial communities and their differential responses to modified gaseous environments. More recently, Tejedor-Calvo et al. (2020) investigated the effects of γ-irradiation at doses ranging from 0.5 to 2.5 kGy on T. aestivum packaged under modified atmosphere. The results showed that γ-irradiation, regardless of dose, effectively improved microbiological stability during refrigerated storage. All treatments caused an immediate reduction of the monitored microbial populations by more than 3 log cfu/g, while higher irradiation doses progressively slowed microbial regrowth throughout the entire storage period (42 days at 4°C). Successively the same research group evaluated a gelatin hydrogel-based packaging (GHP) system to extend the shelf life of fresh T. melanosporum while preserving its aroma profile (Tejedor-Calvo et al., 2024). Compared with microperforated modified atmosphere packaging (MAP) and macroperforated control packaging, GHP maintained a low microbial load throughout storage (<4 log CFU/g after 28 days). Similarly, Daei et al. (2023) demonstrated that mild thermal processing is effective in reducing microbial contamination and extending shelf life; in particular, treatment at 63°C for 3 min resulted in a reduction of 3.05–3.2 log CFU/g of total aerobic bacteria compared to untreated controls. Furthermore, Reale et al. (2009) reported that fresh black truffles are heavily contaminated by diverse microbial groups and showed that irradiation doses above 2 kGy achieved effective microbial decontamination but negatively affected sensory properties, whereas irradiation at 1.5 kGy successfully extended shelf life at 4°C with limited sensory impact. The effects of two doses of electron beam irradiation on the microbial populations (total mesophilic aerobes, Pseudomonas genus, Enterobacteriaceae family, molds and yeasts of T. aestivum packaged under modified atmospheres were monitored during 42 days of storage at 4°C (Rivera et al., 2011a). Irradiation at 1.5 and 2.5 kGy resulted in marked reductions of Pseudomonad populations, corresponding to decreases of 4.3 and 5.5 log units, respectively. Enterobacteriaceae were reduced by 5.4 log units following treatment at 1.5 kGy, while counts fell below the detection limit (<1.0 log cfu/g) at the higher dose. In contrast, lactic acid bacteria and yeasts were less susceptible to ionizing radiation and emerged as the dominant microbial groups during storage, reaching population levels of up to 7.1 log cfu/g.
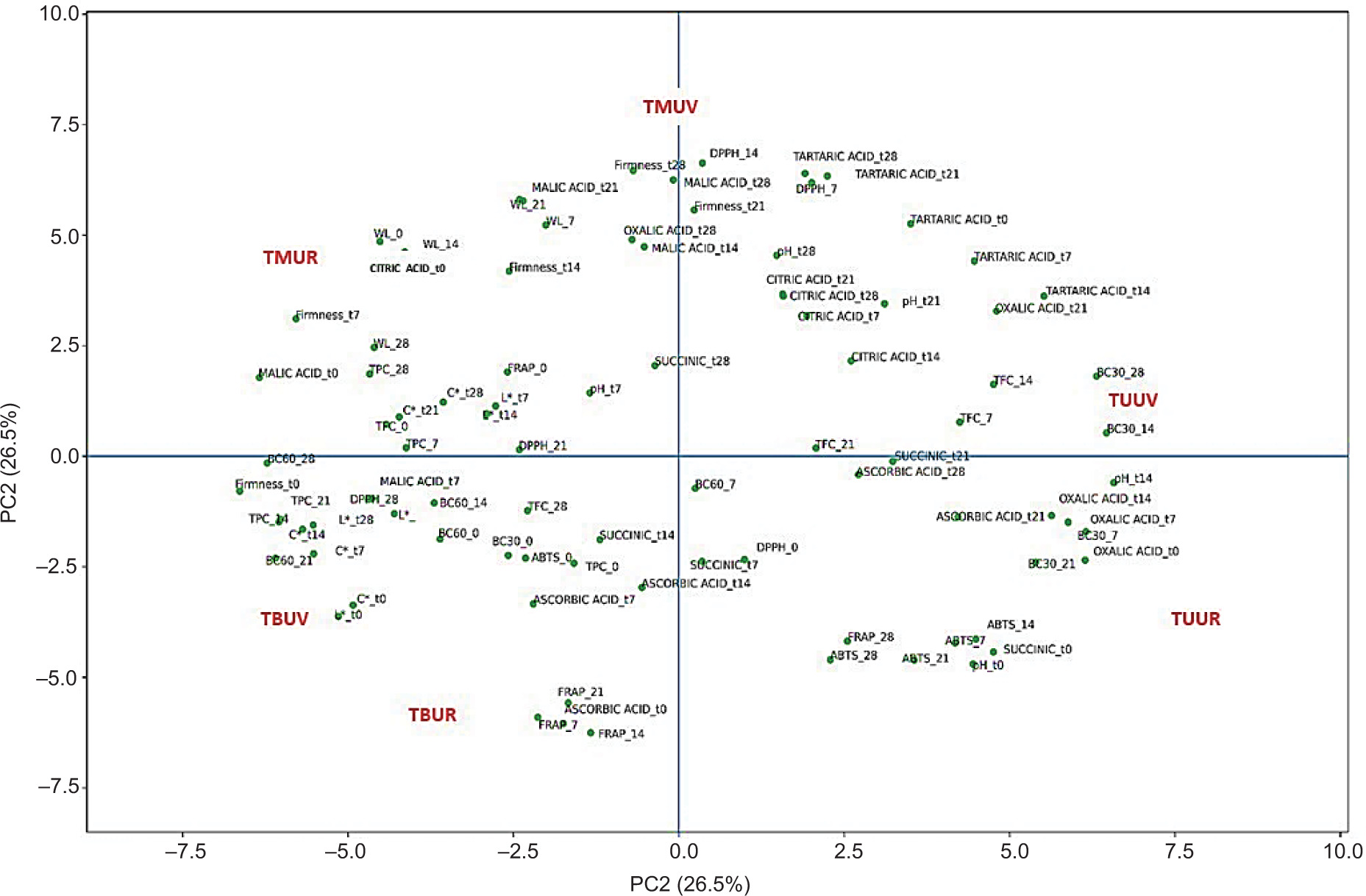
Figure 5. PCA biplot (PC1 vs PC2) showing the simultaneous projection of truffle samples (scores) and quality-related variables (loadings). The plot highlights the separation between samples stored under rice (UR) and under vacuum (UV), as well as the variables contributing to the discrimination.
In conclusion, total microbial load in fresh truffle foods was significantly affected by storage time, preservation method, and species, confirming their high perishability. Total aerobic mesophilic counts proved to be a useful indicator for describing overall microbial evolution and for comparison with previous studies, although they do not capture qualitative shifts in microbial composition. The species-specific responses observed under reduced oxygen conditions further highlight the complexity of truffle-associated microbiota and the need for integrated microbiological approaches in future investigations.
Analysis of organic acid in truffles stored in different conditions
Oxalic, tartaric, malic, citric, succinic, and fumaric acids were quantified in truffles as quality parameters (Table 4). In all truffles, citric acid resulted in the most abundant compound with values of 218.96, 405.27, and 636.95 mg/g extract for TU, TB, and TM, respectively.
Table 4. UHPLC analysis of selected acid markers in truffles subjected to different storage conditions.
| Organic acids (mg/g of truffle extract) | |||||||
|---|---|---|---|---|---|---|---|
| TIME | OXALIC AC. | TARTARIC AC. | MALIC AC. | ASCORBIC AC. | CITRIC AC. | SUCCINIC AC. | |
| TUUV | 0 | 67.00 ± 5.60a | 331.42 ± 11.90a | 217.18 ± 10.00a | 3.51 ± 0.32a | 218.96 ± 10.02e | 88.79 ± 6.80d |
| 7 | 71.43 ± 6.90b | 309.48 ± 11.60b | 185.47 ± 9.80b | 1.59 ± 0.15b | 1443.76 ± 38.80c | 84.00 ± 6.40e | |
| 14 | 54.09 ± 4.80c | 292.88 ± 10.80c | 114.73 ± 9.10c | 1.85 ± 0.18b | 2198.10 ± 40.10b | 97.33 ± 7.70c | |
| 21 | 44.28 ± 4.20d | 143.76 ± 8.80d | 128.77 ± 9.20d | 1.70 ± 0.17b | 952.38 ± 24.01d | 129.86 ± 9.30b | |
| 28 | 36.82 ± 3.80e | 100.49 ± 8.30e | 119.67 ± 9.10e | 1.59 ± 0.15b | 2325.23 ± 40.20a | 131.58 ± 9.50a | |
| Sign. | ** | ** | ** | ** | ** | ** | |
| TUUR | 0 | 67.00 ± 5.60a | 331.42 ± 11.90a | 217.18 ± 10.10a | 3.51 ± 0.35a | 218.96 ± 10.10a | 88.79 ± 6.80d |
| 7 | 63.85 ± 5.40b | 253.88 ± 10.50b | 185.39 ± 9.80b | 0.77 ± 0.07b | 188.83 ± 9.80b | 110.02 ± 9.10a | |
| 14 | 59.47 ± 4.90c | 201.82 ± 10.10c | 137.88 ± 9.30c | 0.66 ± 0.06b | 303.76 ± 11.00c | 104.75 ± 9.00b | |
| 21 | 41.46 ± 4.10d | 73.16 ± 6.90d | 36.83 ± 3.20d | 0.74 ± 0.07b | 785.13 ± 21.80d | 103.02 ± 8.90c | |
| 28 | 8.78 ± 0.80e | 76.68 ± 7.20d | 34.00 ± 3.10e | 0.20 ± 0.02c | 56.20 ± 5.50e | 19.02 ± 1.80e | |
| Sign. | ** | ** | ** | ** | ** | ** | |
| TBUV | 0 | 28.68 ± 2.70a | 177.26 ± 9.40a | 289.58 ± 10.60a | 6.61 ± 0.61a | 405.27 ± 12.10e | 74.19 ± 7.40a |
| 7 | 26.70 ± 2.60b | 116.31 ± 8.60b | 211.89 ± 10.20b | 1.68 ± 0.70b | 468.23 ± 12.60d | 153.05 ± 9.80b | |
| 14 | 16.09 ± 1.50d | 31.15 ± 3.00c | 97.53 ± 8.00c | 1.83 ± 0.80b | 870.83 ± 22.02b | 242.00 ± 10.40c | |
| 21 | 13.78 ± 1.30d | 22.99 ± 2.30d | 86.09 ± 7.90d | 0.73 ± 0.07c | 740.10 ± 21.02c | 116.19 ± 9.60c | |
| 28 | 16.88 ± 1.50c | 10.22 ± 1.02e | 38.11 ± 4.20e | 0.43 ± 0.04d | 1128.47 ± 38.00a | 115.30 ± 9.50d | |
| Sign. | ** | ** | ** | ** | ** | ** | |
| TBUR | 0 | 28.68 ± 2.70 a | 177.26 ± 9.40a | 289.58 ± 10.70a | 6.61 ± 0.66a | 405.27 ± 12.01a | 74.19 ± 7.00a |
| 7 | 19.58 ± 1.90b | 144.03 ± 8.80b | 236.43 ± 10.30b | 1.79 ± 0.17b | 360.32 ± 11.60b | 44.80 ± 4.20b | |
| 14 | 28.63 ± 2.70 a | 80.81 ± 6.20c | 80.61 ± 6.10c | 1.30 ± 0.13b | 479.18 ± 12.70e | 22.10 ± 2.20c | |
| 21 | 29.82 ± 2.90a | 25.63 ± 2.50d | 82.05 ± 6.50c | 0.46 ± 0.04e | 628.97 ± 28.00d | 34.28 ± 3.40d | |
| 28 | 15.89 ± 1.50c | 7.72 ± 0.70e | 42.58 ± 4.20d | 0.64 ± 0.06d | 219.96 ± 10.10c | 13.28 ± 1.30e | |
| Sign. | ** | ** | ** | ** | ** | ** | |
| TMUV | 0 | 19.91 ± 1.90a | 361.02 ± 11.60a | 296.56 ± 10.80a | 1.63 ± 1.20a | 636.95 ± 28.00e | 56.84 ± 5.60e |
| 7 | 14.64 ± 1.40b | 259.89 ± 10.20b | 173.96 ± 9.70b | 1.03 ± 1.00b | 698.07 ± 28.20d | 67.88 ± 6.70d | |
| 14 | 16.71 ± 1.60c | 221.28 ± 10.20c | 154.45 ± 9.50c | 0.65 ± 0.06c | 1176.23 ± 38.40b | 57.57 ± 5.70b | |
| 21 | 38.97 ± 3.90e | 185.42 ± 9.80d | 165.74 ± 9.60d | 0.28 ± 0.02d | 2028.22 ± 40.10c | 68.74 ± 6.80c | |
| 28 | 27.43 ± 2.70d | 177.95 ± 9.70e | 133.78 ± 9.30e | 0.32 ± 0.03c | 2185.34 ± 40.20a | 63.16 ± 6.30a | |
| Sign. | ** | ** | ** | ** | ** | ** | |
| TMUR | 0 | 19.91 ± 1.80a | 361.02 ± 11.60a | 296.56 ± 10.90a | 1.63 ± 1.00a | 636.95 ± 28.00b | 56.84 ± 5.60a |
| 7 | 26.73 ± 2.70b | 273.87 ± 10.70b | 265.07 ± 10.60b | 0.88 ± 0.08b | 686.11 ± 28.20a | 58.33 ± 5.8ab | |
| 14 | 36.62 ± 3.60c | 150.96 ± 9.50c | 227.07 ± 10.20c | 0.67 ± 0.06c | 381.15 ± 11.80c | 71.50 ± 7.10b | |
| 21 | 39.46 ± 3.90d | 131.50 ± 9.30d | 200.120 ± 10.00d | 0.28 ± 0.02d | 266.91 ± 10.30d | 74.66 ± 7.40c | |
| 28 | 40.81 ± 4.00e | 127.33 ± 9.20e | 145.15 ± 9.40e | 0.21 ± 0.02e | 198.76 ± 9.80e | 77.21 ± 7.70c | |
| Sign. | ** | ** | ** | ** | ** | ** | |
TUUR: Tuber aestivum var. uncinatum under rice; TUUV: Tuber aestivum var. uncinatum under vacuum; TBUR: Tuber borchii under rice; TBUV: Tuber borchii under vacuum. TMUR: Tuber mesentericum under rice; TMUV: Tuber mesentericum under vacuum. Data are expressed as means ± SD (n = 3). The differences in the function of storage time (column) were analyzed by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant at *P ≤ 0.05; **P ≤ 0.01.
Tartaric acid and malic acid are also well represented in all fresh truffles. It is interesting to note that both citric and succinic acids significantly increase during storage in all truffles subjected to vacuum storage conditions. Moreover, an increase in malic acids during storage was observed in TM samples.
Previously, Tejedor-Calvo et al. (2021) analysed the organic acid content of different truffle species. Oxalic acid resulted in the most abundant compounds, Terfezia truffles, with values of 141.9 and 37 mg/100 g DW for Tuber arenaria and Tuber magnussi, respectively, whereas fumaric acid was detected only in TM (≈ 0.03 mg/100 g DW).
Evolution of bioactive compounds (TPC and TFC) in truffles in different storage conditions
To evaluate the impact of truffle storage on TPC and TFC, spectrophotometric assays were applied. Fresh ascomycetes extract at Day 0 of storage showed TPC in the range 14.40–16.40 mg chlorogenic acid equivalent (CAE)/g fresh weight (FW) for TU and TM, respectively (Table 5). In all samples, a reduction in TPC was observed, which was more accentuated in samples stored under rice, with a reduction of −74.19 and −86.59% for TB and TU, respectively, after 28 days of storage at 4°C. This reduction is significant starting from Day 7 of storage and may be the consequence of both the metabolic activities of the truffle and the microorganisms it contains.
Table 5. Evolution of TPC (mg CAE/g FW), and TFC (mg QE/g FW) in truffles storage under rice (TUUR, TBUR, TMUR) and under vacuum (TUUV, TBUV, TMUV) for 28 days at 4°C.
| 0 | 7 | 14 | 21 | 28 | Sign. | |
|---|---|---|---|---|---|---|
| TPC mg CAE/g FW | ||||||
| TUUR | 14.40 ± 0.22a | 13.50 ± 1.13b | 8.70 ± 0.81c | 5.60 ± 0.61d | 2.20 ± 0.20e | ** |
| TUUV | 16.40 ± 2.32a | 8.80 ± 0.80b | 7.40 ± 0.75b | 6.70 ± 0.71c | 6.10 ± 0.60c | ** |
| TMUR | 15.55 ± 1.56a | 14.80 ± 1.15a | 11.60 ± 1.12b | 8.80 ± 0.60c | 6.70 ± 0.18d | ** |
| TMUV | 14.40 ± 1.34a | 13.30 ± 1.13a | 9.30 ± 0.81b | 6.40 ± 0.70c | 5.40 ± 0.50d | ** |
| TBUR | 16.40 ± 2.65a | 12.50 ± 1.12b | 11.90 ± 1.12b | 8.80 ± 0.90c | 6.00 ± 0.40d | ** |
| TBUV | 15.55 ± 2.02a | 14.30 ± 1.14a | 10.80 ± 0.70b | 8.50 ± 0.60c | 6.20 ± 0.40d | ** |
| TFC (mg QE/g FW) | ||||||
| TUUR | 5.33 ± 0.66b | 5.80 ± 0.56a | 4.30 ± 0.40c | 3.30 ± 0.31d | 2.30 ± 0.21e | ** |
| TUUV | 5.12 ± 0.45b | 5.90 ± 0.60a | 4.70 ± 0.50c | 3.50 ± 0.34d | 2.00 ± 0.20e | ** |
| TMUR | 6.24 ± 0.68a | 4.70 ± 0.52b | 3.50 ± 0.36c | 2.40 ± 0.22d | 1.70 ± 0.16e | ** |
| TMUV | 5.33 ± 0.23b | 5.70 ± 0.55a | 4.70 ± 0.40c | 4.00 ± 0.39d | 3.10 ± 0.30e | ** |
| TBUR | 5.12 ± 0.78a | 4.60 ± 0.51b | 4.10 ± 0.38c | 3.60 ± 0.35d | 2.90 ± 0.28e | ** |
| TBUV | 6.24 ± 0.61a | 5.70 ± 0.55b | 3.80 ± 0.39c | 3.20 ± 0.33d | 3.00 ± 0.30d | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TUUV: Tuber aestivum var. uncinatum under vacuum; TBUR: Tuber borchii under rice; TBUV: Tuber borchii under vacuum. TMUR: Tuber mesentericum under rice; TMUV: Tuber mesentericum under vacuum. Data are expressed as means ± SD (n = 3). The differences in the function of storage time (row) were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant at *P ≤ 0.05; **P ≤ 0.01. n.s.: not significant.
A similar state was observed in TFC, where a maximum reduction was observed after 28 days of storage in TMUR samples with a value of −70.18%. On the contrary, the same truffles stored under vacuum showed a reduction in TFC by −47.46%. This could be related to the lack of oxygen in the samples stored under vacuum, which requires the functioning of enzymes involved in the metabolic processes of these phytochemicals, such as polyphenol oxidase (PPO). This enzyme is responsible for the oxidation of phenols that has as consequences the reduction in TPC (Sicari et al., 2024).
Previously, Savini et al. (2017) investigated the effect of the storage conditions for 28 days on T. melanosporum. For this purpose, samples were refrigerated at 4°C under different packaging conditions: air atmosphere (A), reduced pressure (V), mix 1% O2/99% N2 (ON) and mix 40% CO2/60% N2 (CN). A decrease in TPC content was observed during storage regardless of the packaging adopted. Sample ON showed the highest TPC with a value of 33.3 GAE/g dried extract. T. aestivum, T. -brumale, Tuber indicum, Tuber gennadii, T. magnatum, T. melanosporum, and Tuber oligospermum, Terfezia arenaria, Terfezia leptoderma, and Terfezia magnusii were screened for their TPC. T. magnatum was the species with the highest TPC values (290 mg GAE/100 g), followed by T. melanosporum and T. leptoderma (138 and 134 mg GAE/100 g, respectively). In contrast, T. indicum species obtained the minor levels (22 mg GAE/100 g) (Tejedor-Calvo et al., 2021). The TPC in Chinese edible truffles, Tuber latisporum, Tuber subglobosum and Tuber pseudohimalayense were evaluated. T. pseudohimalayense’s methanol extract showed the highest TPC (735.01 mg GAE/100 g DW), whereas T. subglobosum was characterized by the highest TFC (1355.43 mg rutin/100 g DW) (Yan et al., 2017). Previously, Hajjar et al. (2010) investigated the effect of storage in high CO2 and low O2 atmospheres on TU truffles and found that the truffle’s TPC significantly decreased after 16 days of storage under CO2, while it remained stable for 14 days in air-stored truffles. At the end of the storage period, samples kept in air had a significantly higher TPC than truffles stored under CO2, with values of 27.9 and 23.2 mg GAE/mL, respectively. A high content of phenolic compounds was also found in Reddellomyces westraliensis methanol extract (18.94 μg protocatechuic equivalents/mg extract) (Çayan et al., 2022). A positive effect on TPC was obtained by Tejedor-Calvo et al. (2020) who applied γ-irradiation on T. aestivum packaged under modified atmosphere. In fact, these compounds increased during storage, reaching approximately twice the initial levels. Alothman et al. (2009) suggested that irradiation may disrupt the chemical linkages that bind phenolic compounds to other molecular structures, leading to the release of phenols in their soluble form. In contrast, Altamim et al. (2024) evidenced that γ-irradiation significantly affected the total phenolic (TPC) and total flavonoid (TFC) contents of Zubaidi (Terfezia boudieri), showing a progressive decrease in both parameters with increasing dose.
A kinetic model was applied to determine the change in product quality due to different treatments of truffles. In the experiment, the degradation kinetics of quantified bioactive compounds (TPC and TFC) were studied. The data show that all the analysed parameters follow a zero-order reaction as reported in Table S3. Analysis of results showed in general that the t1/2 trend for both TPC and TFC depends not on the different conservation method but on the truffle species analysed. For TPC, the lowest t1/2 value was found for TUUR (1.5020 days), whereas the same truffle storage under vacuum (TUUV) exhibited the highest t1/2 value (2.1369 days). For the TFC sample, TMUR showed the lowest half-life time with a t1/2 value of 4.2620 days, whereas TBUR demonstrated the highest value (8.9189 days). During storage, the reduction of both TPC and TFC is probably due to the activity of polyphenol oxidase (PPO), the enzyme responsible for the oxidation of phenols that is present in truffles (Benaceur et al., 2020).
Antioxidant activity
The antioxidant activity of truffle samples stored under different conditions was investigated, and the results are reported in Tables 6–8. Overall, samples exhibited ABTS radical scavenging activity in the range of 251.52–258.76 µg/mL for TU and TM, respectively, whereas higher IC50 values (316.72–391.50 µg/mL for TU and TM, respectively) were recorded in the DPPH assay at Day 0 (Table 6). For both assays, a progressive reduction in antioxidant activity was observed over the storage period. After 28 days, TBUV showed the lowest IC50 value (283.00 µg/mL), followed by TUUV (284.77 µg/mL). Among truffles stored under rice, TM exhibited the highest antioxidant potential against the ABTS radical, with an IC50 value of 296.10 µg/mL. A similar decrease in DPPH radical scavenging activity was observed during storage, with IC50 values ranging from 426.68 to 499.62 µg/mL for TUUR and TBUR, respectively. A slight reduction in IC50 values was recorded over time in the β-carotene bleaching test, regardless of the storage conditions (Table 7). After 28 days of storage at 4°C, IC50 values ranged from 21.33 to 24.52 µg/mL and from 25.28 to 26.50 µg/mL for TU and TB after 30 and 60 minutes of incubation, respectively. At the end of the storage period, TBUV exhibited the highest protection against lipid peroxidation after 30 min of incubation (IC50 = 27.42 µg/mL), followed by TMUR, whereas TUUR showed the highest activity after 60 min. In the FRAP assay, values ranged from 26.12 to 36.13 µM Fe(II)/g for TM and TB, respectively. Although these values were approximately one-third lower than those of the BHT positive control (Table 8), a reduction in ferric reducing antioxidant power was observed over time, with values decreasing to 12.15–22.90 µM Fe(II)/g for TMUV and TUV, respectively.
Table 6. Evolution of radical scavenging activities, evaluated by ABTS and DPPH tests, in truffles during storage.
| Samples | Days storage | |||||
|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 21 | 28 | Sign | |
| ABTS (IC50 µg/mL) | ||||||
| TUUR | 251.50 ± 5.22e | 303.12 ± 4.60d | 359.93 ± 8.40c | 390.62 ± 9.40b | 393.99 ± 6.80a | ** |
| TUUV | 258.70 ± 4.76e | 292.50 ± 8.60d | 298.00 ± 8.90c | 302.21± 9.00b | 308.22 ± 9.60a | ** |
| TMUR | 256.70 ± 3.89e | 266.72 ± 7.30d | 270.90 ± 8.40cd | 281.21 ± 8.50c | 296.10 ± 7.80a | ** |
| TMUV | 251.50 ± 5.21e | 258.92 ± 8.70d | 262.73 ± 6.50cd | 270.41 ± 6.80c | 284.77 ± 6.10a | ** |
| TBUR | 258.70 ± 4.23e | 299.83 ± 5.90d | 315.85 ± 7.70c | 351.12 ± 6.40b | 399.47 ± 7.50a | ** |
| TBUV | 256.70 ± 4.54d | 261.28 ± 6.00cd | 267.35 ± 7.40c | 280.70 ± 3.70b | 283.00 ± 5.70a | ** |
| DPPH (IC50 µg/mL) | ||||||
| TUUR | 316.70 ± 5.23e | 368.11 ± 6.30d | 373.52 ± 7.20c | 400.71 ± 7.40b | 426.68 ± 8.10a | ** |
| TUUV | 391.50 ± 5.65d | 381.80 ± 4.90d | 398.92 ± 6.60c | 412.31± 7.30b | 434.48 ± 7.50a | ** |
| TMUR | 322.60 ± 4.22e | 392.31 ± 8.70d | 400.70 ± 7.90c | 423.32 ± 8.40b | 477.61 ± 12.00a | ** |
| TMUV | 316.70 ± 5.72e | 398.66 ± 6.50d | 419.00 ± 7.50c | 428.13 ± 9.10b | 445.91 ± 6.90a | ** |
| TBUR | 391.50 ± 4.78d | 350.00 ± 5.70e | 372.62 ± 7.30d | 456.82 ± 11.00b | 499.62 ± 10.70a | ** |
| TBUV | 322.60 ± 4.54d | 340.82 ± 7.80d | 374.81 ± 9.10c | 398.20 ± 8.40b | 445.30 ± 6.60a | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TUUV: Tuber aestivum var. uncinatum under vacuum; TBUR: Tuber borchii under rice; TBUV: Tuber borchii under vacuum. TMUR: Tuber mesentericum under rice; TMUV: Tuber mesentericum under vacuum. Data are expressed as means ± SD (n = 3). Ascorbic acid was used as positive control with IC50 values of 1.78 ± 0.29 and 5.11 ± 0.94 µg/mL in ABTS and DPPH test, respectively. The differences in the function of storage time (row) were evaluated by one-way ANOVA followed by Tukey’s post-hoc test. Results followed by letters are significant. *P ≤ 0.05; **P ≤ 0.01.
Table 7. Evolution of protection from lipid peroxidation evaluated by ß-carotene bleaching test in truffles during storage.
| Samples | Days storage | |||||
|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 21 | 28 | ||
| β-carotene bleaching test at 30 min incubation (IC50 µg/mL) | ||||||
| TUUR | 21.30 ± 1.55a | 28.90 ± 2.20b | 29.77 ± 1.90bc | 31.87 ± 2.30cd | 33.27 ± 3.90d | ** |
| TUUV | 24.50 ± 1.34a | 27.91 ± 2.40b | 28.97 ± 2.60bc | 29.27 ± 3.00c | 33.57 ± 1.80d | ** |
| TMUR | 23.80 ± 2.04a | 25.82 ± 2.70ab | 26.99 ± 2.10ab | 27.72 ± 3.90c | 27.99 ± 4.30d | ** |
| TMUV | 21.30 ± 2.05a | 25.78 ± 2.10b | 28.15 ± 2.30c | 26.29 ± 2.40b | 32.50 ± 3.10d | ** |
| TBUR | 24.50 ± 2.11a | 25.53 ± 3.40a | 26.56 ± 2.70ab | 26.92 ± 2.40b | 28.30 ± 3.10c | ** |
| TBUV | 23.80 ± 1.98a | 26.14 ± 1.90ab | 27.18 ± 2.80b | 27.33 ± 2.50b | 27.91 ± 2.80c | ** |
| β-carotene bleaching test at 60 min incubation (IC50 µg/mL) | ||||||
| TUUR | 25.20 ± 1.23a | 25.90 ± 2.30a | 26.51 ± 2.60ab | 27.82 ± 2.60b | 29.22 ± 4.20c | ** |
| TUUV | 26.40 ± 2.04a | 26.80 ± 2.60 a | 27.42 ± 3.44ab | 28.22 ± 2.50b | 30.10 ± 3.80c | ** |
| TMUR | 26.50 ± 2.42a | 26.70 ± 2.50 a | 29.92 ± 3.31b | 33.66 ± 3.70c | 37.90 ± 3.90d | ** |
| TMUV | 25.20 ± 2.52a | 25.52 ± 2.33 a | 27.82 ± 2.10ab | 28.41 ± 2.70b | 33.33 ± 5.00c | ** |
| TBUR | 26.40 ± 1.36a | 26.41 ± 2.71a | 32.21 ± 2.32b | 34.22 ± 3.10c | 37.80 ± 4.20d | ** |
| TBUV | 26.50 ± 1.85a | 25.91 ± 2.60a | 27.10 ± 2.71ab | 33.79 ± 3.60c | 34.90 ± 3.80c | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TUUV: Tuber aestivum var. uncinatum under vacuum; TBUR: Tuber borchii under rice; TBUV: Tuber borchii under vacuum. TMUR: Tuber mesentericum under rice; TMUV: Tuber mesentericum under vacuum. Data are expressed as means ± SD (n = 3). Propyl gallate was used as positive control with IC50 values of 0.11 ± 0.06 and 0.9 ± 0.04 µg/mL in ß-carotene bleaching test after 30- and 60-min incubation, respectively. The differences in the function of storage time (row) were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant. *P ≤ 0.05; **P ≤ 0.01. n.s.: not significant.
Table 8. Evolution of ferric reducing ability powder (FRAP) in truffles during storage.
| Samples | Days storage | Sign. | ||||
|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 21 | 28 | ||
| FRAP values (μM Fe (II)/g) | ||||||
| TUUR | 33.20 ± 1.23a | 32.33 ± 2.90a | 27.01 ± 2.10b | 21.55 ± 1.60c | 20.10 ± 1.30c | ** |
| TUUV | 26.1 ± 1.440a | 25.32 ± 1.80ab | 23.93 ± 2.00bc | 23.31 ± 1.70c | 22.90 ± 1.50c | ** |
| TMUR | 36.10 ± 2.03a | 25.41 ± 1.80b | 20.19 ± 1.70c | 18.99 ± 1.30c | 12.15 ± 1.20d | ** |
| TMUV | 33.20 ± 1.77a | 25.31 ± 1.60b | 20.18 ± 1.70c | 14.20 ± 1.20d | 12.15 ± 1.20e | ** |
| TBUR | 36.1 ± 2.020a | 33.88 ± 1.70b | 30.38 ± 1.60c | 25.20 ± 1.90d | 16.22 ± 1.40d | ** |
| TBUV | 36.10 ± 2.32a | 35.18 ± 3.80a | 32.20 ± 3.10b | 30.28 ± 2.30b | 22.90 ± 1.50e | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TUUV: Tuber aestivum var. uncinatum under vacuum; TBUR: Tuber borchii under rice; TBUV: Tuber borchii under vacuum. TMUR: Tuber mesentericum under rice; TMUV: Tuber mesentericum under vacuum. Data are expressed as means ± SD (n = 3). BHT was used as positive control with IC50 value of 63.45 ± 5.2 μMFe (II)/g. The differences in the function of storage time (row) were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant. *P ≤ 0.05; **P ≤ 0.01.
Overall, these results do not allow the identification of a clear trend or a single preservation method capable of effectively maintaining the antioxidant capacity of truffles. This behavior is likely related to the fact that antioxidant activity is not attributable to a single compound or class of compounds, but rather to the synergistic interactions among the various phytochemicals present in the extracts (Bayram et al., 2023). Previous studies support this interpretation. Savini et al. (2017) demonstrated that the antioxidant activity of T. melanosporum truffles decreases markedly over time, regardless of the packaging system used, although vacuum storage preserved radical scavenging activity more effectively for up to 14 days. After this period, the highest ABTS radical scavenging activity was observed in samples stored under modified atmosphere packaging (MAP; 40% CO2/60% N2), with an IC50 value of 60.0 µg/mL. Conversely, in the DPPH assay, the best antioxidant performance was maintained for up to 21 days in vacuum-stored samples (IC50 values of 55.7 and 84.5 µg/mL after 7 and 21 days, respectively). In contrast to our findings, Savini et al. (2017) reported consistently high FRAP values (104.4–128.4 mM Fe(II)/g), irrespective of the packaging system. Çayan et al. (2022) investigated the antioxidant activity of different solvent extracts (acetone, chloroform, methanol, n-hexane, and water) from Reddellomyces westraliensis. Except for the aqueous extract (IC50 = 172.40 µg/mL), none of the extracts showed activity against the DPPH radical. In contrast, ABTS IC50 values ranged from 15.98 to 196.69 µg/mL for the aqueous and chloroform extracts, respectively. The highest protection against lipid peroxidation was observed for the acetone extract (IC50 = 5.66 µg/mL), a value markedly lower than those recorded for our -samples at time 0.
Tejedor-Calvo et al. (2021) reported DPPH radical scavenging values of 4.3 and 3.9 mmol TE/100 g FW for T. magnatum and T. indicum, respectively, while T. gennadii exhibited the lowest activity (0.18 mmol TE/100 g FW). These results are consistent with those reported by Guo et al. (2011), who observed approximately 98% radical reduction for various T. indicum extracts tested at a concentration of 30 mg/mL. In the FRAP assay, T. magnatum and T. indicum again showed the highest antioxidant capacity (5.6–8.8 mmol TE/100 g FW). For Terfezia species, FRAP values ranged from 2.6 to 5.3 mmol TE/100 g FW for T. arenaria and T. leptoderma, respectively (Tejedor-Calvo et al., 2021), in agreement with values reported for other species of the same genus (Al-Laith et al., 2010). Comparable DPPH radical scavenging activity to that observed in our study was reported by Yan et al. (2017) for fresh Chinese truffles, with the following activity ranking: Tuber pseudohimalayense > Tuber latisporum > Tuber subglobosum. Similarly, Çayan et al. (2022) found that the aqueous extract of R. westraliensis exhibited the highest activity in both ABTS and DPPH assays, whereas the acetone extract was the most effective in the β-carotene bleaching test. Results of radical scavenging activity on γ-irradiated Terfezia boudieri evidenced that despite the reduction in TPC and TFC, the DPPH racìdical scavenging effect shows a dose--dependent increase, reaching its maximum value at 10 kGy. This apparent paradox indicates that the antioxidant activity does not depend exclusively on the total amount of phenols and flavonoids, but also on the formation of new antioxidant compounds or on a greater availability of highly reactive radical scavenging fractions generated during irradiation (Altamim, 2024).
Sensory analysis
The analysis of the sensory parameters (Table 9) highlights values between 8.70 and 6.80 for TM and TB, respectively, on Day 0. In general, a significant variation in texture was observed in the samples preserved under rice with reference to TBUR (5.10). Regarding aroma parameter values in the range 4.80–5.00 were found from fresh truffles. With a “fully typical” descriptor reported by the panelist only for the TM sample. Also in this case, as per other literature works (Rivera et al. 2010), conservation over time causes a loss of aroma with better performances for vacuum conservation with values between 3.30 and 3.50 for TBUV and TUUV, respectively, after 28 days of storage. Aroma values of 5.0 and 1.0 were reported for TM and TU storage, respectively, under air after 28 days of storage, whereas values of 7.2 and 5.8 were found for the same sample storage under MAP conditions (Rivera et al., 2010). The perception of the flavor descriptor follows the progress of the aroma, with the lowest values after 28 days of storage in TM and TB. Regarding the Global Judgement, no truffle at the end of the conservation period is unacceptable. Savini et al. (2020) examined hypobaric packaging at 30 kPa with 1% O2/99% N2 and modified atmosphere packaging (40% CO2/60% N2) to extend the shelf life of black truffles stored at 4°C in sealed polypropylene containers. Texture remained unchanged after 7 days, while differences among MAP treatments emerged after 28 days. Although texture was preserved up to day 28, aroma degradation occurred regardless of packaging conditions. In the present study, both texture and aroma progressively deteriorated during storage. Likewise, Amadoro et al. (2013) reported a substantial decline in sensory quality, with overall scores decreasing from 15 at Day 0 to 4.3 at Day 28; treatments based on chitosan, ultrasound, and modified atmosphere packaging, applied alone or in combination, resulted in intermediate scores of approximately 6.
Table 9. Sensory parameters (texture, aroma, flavor, and global judgement) of truffles at Day 0 and at 7, 14, 21, and 28 days of storage at 4 °C under rice and under vacuum.
| Days storage (die) | 0 | 7 | 14 | 21 | 28 |
|---|---|---|---|---|---|
| Median of parameter texture1 | |||||
| TUUR | 7.3 | 7.9 | 7.1 | 6.6 | 6.0 |
| TUUV | 7.3 | 6.4 | 5.9 | 5.6 | 5.2 |
| TMUR | 8.7 | 7.5 | 7.4 | 7.0 | 6.5 |
| TMUV | 8.7 | 8.3 | 8.2 | 8.0 | 7.2 |
| TBUR | 6.8 | 6.1 | 6.0 | 5.9 | 5.1 |
| TBUV | 6.8 | 6.6 | 6.4 | 6.2 | 6.0 |
| Median of parameter aroma2 | |||||
| TUUR | 4.8 | 4.6 | 4.2 | 3.5 | 3.1 |
| TUUV | 4.8 | 4.6 | 4.4 | 3.9 | 3.5 |
| TMUR | 5.0 | 4.7 | 4.1 | 3.4 | 3.2 |
| TMUV | 5.0 | 4.7 | 4.4 | 3.9 | 3.4 |
| TBUR | 4.9 | 4.4 | 4.0 | 3.7 | 3.0 |
| TBUV | 4.9 | 4.3 | 4.1 | 3.8 | 3.3 |
| Median of parameter flavour3 | |||||
| TUUR | 4.2 | 4.0 | 3.8 | 3.5 | 3.0 |
| TUUV | 4.2 | 4.2 | 3.8 | 3.8 | 3.5 |
| TMUR | 4.0 | 3.8 | 3.6 | 3.4 | 2.8 |
| TMUV | 4.0 | 4.0 | 3.8 | 3.8 | 3.6 |
| TBUR | 4.0 | 3.9 | 3.6 | 3.3 | 2.8 |
| TBUV | 4.0 | 4.0 | 3.8 | 3.5 | 3.2 |
| Median of parameter global Judgement4 | |||||
| TUUR | 5.0 | 4.9 | 4.2 | 3.8 | 3.7 |
| TUUV | 5.0 | 4.9 | 4.6 | 4.5 | 4.0 |
| TMUR | 5.0 | 4.7 | 4.4 | 3.9 | 3.1 |
| TMUV | 5.0 | 5.0 | 4.6 | 4.1 | 3.7 |
| TBUR | 5.0 | 4.8 | 4.7 | 4.0 | 3.3 |
| TBUV | 5.0 | 4.8 | 4.4 | 3.9 | 3.2 |
TBUR: Tuber borchii Vittad. storage under rice; TMUR: Tuber mesentericum storage under rice; TUUR: Tuber aestivum var. uncinatum storage under rice; TBUV: Tuber borchii Vittad. storage under vacuum; TMUV: TMUR: Tuber mesentericum storage under vacuum; TUUV: Tuber aestivum var. uncinatum storage under vacuum. 1Texture score on a 1–9 scale (1 = unacceptable; 3 = soft; 5 = moderately soft; 7 = moderately hard; 9 = hard); 2Aroma score on a 1–5 scale (1 = no typical; 3 = moderate; 5 = full typical); 3Flavour score on a 1–5 scale (1 = no typical; 3 = moderate; 5 = full typical). 4Global judgment scored on a 1–5 scale (1 = unacceptable; 3 = moderately acceptable; 5 = excellent).
With respect to γ-irradiation, truffles treated with 1.5 kGy maintained acceptable sensory quality for up to 5 weeks. However, sensory scores approached the acceptability limit with the appearance of yeast colonies on the surface, thus limiting shelf life to 35 days. In contrast, truffles treated with 2.5 kGy resulted in the longest preservation of truffle quality, with sensory attributes remaining stable until Day 42 and only limited development of mycelium, yeasts, and prokaryotic microorganisms observed at the end of storage. This improved stability at higher doses may be associated with the composition of the truffle peridium, which is mainly composed of chitin and is therefore susceptible to colonization by chitinase-producing fungi such as Bionectria ichroleuca (Gan et al., 2007). Probably, the lethal and sublethal effects of irradiation on molds and yeasts likely delayed their development for up to seven weeks, in agreement with previous findings reported for T. aestivum treated with e-beam irradiation (Rivera et al., 2011a). In this case, the irradiated batches showed similar sensorial characteristics during the first three weeks of the experiment. However, better scores were obtained for truffles irradiated with 2.5 kGy from Day 28 onwards. It has already been observed that modified atmospheres substantially inhibit the development of surface molds, and that this helps to conserve the aroma and flavor of summer truffles (Rivera et al., 2010). Moreover, truffles irradiated with 2.5 kGy maintained their organoleptic properties to a greater extent, and neither mycelium growth nor development of yeast colonies was observed. This enabled their shelf life to reach 42 days. Internal color changes observed in all batches were due to the normal senescence process.
Previously, Cullere et al. (2012) investigated the effect of electron-beam and γ-irradiation on the volatile aroma profile of T. aestivum and T. melanosporum. Irradiation-induced modifications in the aromatic composition of both species; however, these changes were not perceived in sensory evaluations. In particular, T. melanosporum, electron-beam treatment caused the most pronounced alterations in the volatile profile, whereas γ-irradiation did not result in significant differences compared to non-irradiated samples. Conversely, in T. aestivum, γ-irradiation produced the greatest changes relative to controls, particularly at the 1.5 kGy dose. The observed variations are consistent with irradiation-induced oxidative degradation of fatty acids, as reflected by increased levels of specific carbonyl compounds, including hexanal in T. melanosporum and hexanal, E, E-2,4-nonadienal, and nonanal in T. aestivum. Overall, although irradiation affected the volatile composition in a species- and treatment-dependent manner, the magnitude of these changes was insufficient to compromise sensory quality.
Multivariate evidence and implications for conservation strategies
Multivariate analysis
A multivariate statistical analysis was performed on the effects of individual species, storage system, storage time and their interactions on the main chemical–physical parameters monitored during the 4°C shelf life of truffles. Table 10 shows the results of the multivariate analysis, which clearly indicate that variables cannot be evaluated in isolation. It was highlighted that the sample explained a species-specific “fingerprint” with a significant relevance (P < 0.01), substantially for all parameters, including color parameters, texture, organic acids, weight loss, pH, TBC, TPC and antioxidant tests except for TFC. The Tuber species analyzed therefore, differ in composition and in the biochemical and interspecific changes that persist during the different storage conditions, strongly influenced by intrinsic species characteristics. Time is the factor that most impacts the stability of qualitative characteristics, which inevitably influences the parameters considered due to the biochemical senescence processes that truffles undergo. Packaging, on the other hand, showed a significant effect on some traits, such as C*, firmness and some organic acids and TFC, ABTS, and DPPH assays. Packaging, however, proved to be meaningless for L*, malic acid, pH, TPC, ß-carotene bleaching test at 30 and 60 min and FRAP assay. This test indicated that it significantly affected color saturation rather than luminance, most organic acids (P < 0.01), and oxalic acid (P < 0.05), while no effect was detected on malic acid. The main effect of packing on firmness indicates that the vacuum relative to rice modulates structural integrity,
Table 10. Multivariate statistical analysis of truffle qualities.
| Dependent variable | C* | L* | Firmness | Oxalic acid | Tartaric acid | Malic acid | Ascorbic acid | Citric acid | Succinic acid | Weight loss | pH | TBC | TPC | TFC | ABTS | DPPH | ßcb 30 | ßcb 60 | FRAP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ns | ** | ** | ** | ** | ** |
| Packaging | ** | ns | ** | * | ** | ns | ** | ** | ** | * | ns | ** | ns | ** | ** | * | ns | ns | ns |
| Time | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Sample*Packaging | ** | ** | ns | ** | ** | ** | ** | ** | ** | ns | ns | ** | * | * | ** | ** | ns | ns | ** |
| Sample*Time | ** | ns | ns | ** | ** | ** | ** | ** | ** | * | ** | ** | ns | * | ** | ** | ns | ns | ** |
| Packaging*Time | ** | * | ns | ** | ** | ** | ns | ** | ** | ** | ns | ** | * | ns | ** | ** | ns | ns | ** |
| Sample*Packaging*Time | ns | ns | ns | ** | ** | ** | ns | ** | ** | ns | ** | ** | ** | ** | ** | ** | ns | ns | ns |
ßcb 30: ß-carotene bleaching test at 30 min; ßcb 60: ß-carotene bleaching test at 60 min.
Significance: n.s. not significant; ** Significance at p<0.01; * Significance at p<0.05.
The sample × packaging interaction was highly significant for color characteristics, organic acids, TBC, radical scavenging assays (P < 0.01), significant (P < 0.05) for TPC and TFC, but not for the other parameters analysed, showing how this interaction has different effects for each truffle sample. The sample × time combined effect was not significant for L*, firmness, TPC, ß-carotene bleaching tests, indicating that these changes proceed similarly across species. Not significantly, firmness, ascorbic acid, pH, TFC and ß-carotene bleaching tests showed that differences between packaging are stable during storage time. The three-way interaction (Sample × Packaging × Time) was highly significant (P < 0.01) for organic acids (except for ascorbic acid), pH, TBC, TPC, TFC, and ABTS and DPPH assays. This interaction indicates that the effects of the considered variables and the packaging strategies are not unique but are species-dependent. Overall, it emerged that the two packaging methods have different effects on the parameters considered. In particular, this foundation is highlighted by the strong three-way interaction between organic acids and antioxidant tests. Conservation performance is influenced by the interaction between the species, conservation conditions, and time.
Principal component analysis
Principal Component Analysis (PCA) was applied to explore the relationships among physicochemical and biochemical variables and to evaluate the effect of the different storage conditions on sample discrimination. The first two principal components accounted for 78.8% of the total variance, with PC1 explaining 52.26% and PC2 26.5%, indicating that the bidimensional score plot provides a reliable representation of the dataset (Figure 5).
The PC1 axis was the main source of variability and clearly separated samples according to their overall chemical–metabolic profile versus structural–-physical attributes. Positive PC1 values were strongly associated with organic acids, particularly tartaric, citric and oxalic acids measured at different storage times, as well as with pH. In the lower-right portion of the plot, PC1 was also influenced by variables related to antioxidant activity, including ABTS, FRAP, and succinic acid, suggesting a metabolically active profile. In contrast, negative PC1 values were mainly related to firmness, weight loss (WL), and, to a lesser extent, TPC and some measurements of ascorbic acid, indicating a profile dominated by physical and structural characteristics.
The PC2 axis further differentiated the samples according to the relative contribution of acidity-related variables and antioxidant response. Positive PC2 values were primarily driven by organic acids, particularly tartaric and citric acids, together with some DPPH measurements, whereas negative PC2 values were clearly associated with ABTS and FRAP assays and succinic acid, highlighting a coherent cluster of variables related to antioxidant activity. This distribution suggests that PC2 describes a gradient ranging from an acid-dominated profile to an antioxidant-oriented metabolic response.
The analysis of the scores plot revealed a clear effect of storage conditions within the different sample groups. The TU samples were positioned on the positive side of PC1, indicating a strong association with organic acids and pH. However, a distinct separation between the two storage methods was observed. TUUV (vacuum storage) was located close to the PC1 axis and slightly positive along PC2, suggesting a profile mainly characterized by organic acids and acidity-related variables. Conversely, TUUR (storage under rice) was shifted toward the lower-right quadrant, in proximity to ABTS, FRAP, and succinic acid, indicating a stronger association with antioxidant activity rather than with acidity alone.
In contrast, the TB samples were clustered in the lower left quadrant (negative PC1 and PC2), showing a profile mainly related to TPC, ascorbic acid, and other variables contributing to the negative side of both components. In this group, the separation between TBUR and TBUV was less pronounced, suggesting that, for these samples, the effect of storage method was less influential than the intrinsic variability of the biochemical composition.
A more marked differentiation between storage conditions was observed for the TM samples. TMUR was located in the upper-left quadrant, closely associated with firmness and weight loss, indicating a stronger contribution of structural and physical attributes in samples stored under rice. In contrast, TMUV was positioned at high positive PC2 values near the PC1 origin, suggesting a shift toward a chemically driven profile, mainly influenced by organic acids, and a reduced association with physical parameters.
Overall, the PCA highlights that the impact of storage conditions is strongly dependent on the sample group. Storage under rice (UR) appears to promote, particularly in TU samples, a profile associated with enhanced antioxidant activity, whereas vacuum storage (UV) tends to preserve acidity-related characteristics and, in TM samples, reduces the influence of structural degradation parameters. These findings confirm the usefulness of multivariate analysis in capturing complex and interconnected changes induced by different storage strategies, which may not be evident through univariate analysis alone.
Conclusion
This study provides a comprehensive multidisciplinary evaluation of the taxonomic identity, postharvest quality, and shelf life of three truffle species harvested in Calabria (TM, TB, and TU). The integration of morphological, histological, and molecular analyses ensured accurate species identification, contributing to biodiversity assessment and strengthening traceability within the truffle supply chain.
The postharvest storage trials revealed a pronounced species-specific response to conservation strategies. Among the species investigated, TB showed the highest overall stability, with limited physical, microbiological, and sensory deterioration, particularly under vacuum packaging. Tuber aestivum var. uncinatum exhibited an intermediate behavior, moderately benefiting from vacuum conditions, especially in terms of color and sensory quality. In contrast, TM proved to be the most sensitive species, displaying increased microbial proliferation when stored under vacuum.
With respect to storage methods, vacuum packaging generally slowed firmness loss and color degradation, proving more effective for TB and TU. However, it cannot be considered a universally optimal solution. For TM, traditional rice-based storage ensured better microbiological stability. In addition, this method offers a practical and sustainable advantage, allowing the rice to be recovered and reused as a flavored product.
Despite these differences, neither storage method was able to significantly extend shelf life beyond 21–28 days at 4°C. While both approaches effectively limited weight loss, key quality attributes, including firmness, color, aroma, flavor, and bioactive compound content, progressively declined, with marked deterioration occurring after approximately two weeks. These results confirm the highly perishable nature of fresh truffles.
Microbiological, organic acid, and sensory analyses highlighted ongoing metabolic and microbial activity during storage, influenced by both species and the packaging system. Multivariate statistical analysis demonstrated that species identity exerted a highly significant effect on almost all monitored parameters, defining clear species-specific compositional and metabolic “fingerprints.” Storage time emerged as the main driver of quality decline, reflecting progressive biochemical senescence, whereas packaging showed a selective and parameter-dependent influence, significantly affecting color saturation, firmness, several organic acids, and radical scavenging activity, but exerting limited effects on luminance, pH, and TPC. Notably, the significant Sample × Packaging × Time interaction confirmed that preservation performance depends on the combined and species-dependent interplay between intrinsic biological traits, storage system, and duration. PCA highlighted that rice storage was associated, in some species, with antioxidant-related variables, whereas vacuum packaging tended to preserve acidity-related characteristics and structural integrity, particularly in TM. Sensory evaluation and multivariate outcomes collectively confirmed that quality evolution is driven primarily by intrinsic species characteristics rather than by storage technology alone.
Overall, the findings indicate that postharvest conservation strategies for truffles should be species-specific and tailored to the biological characteristics of each taxon. While vacuum packaging may represent a suitable option for short-term storage under strict temperature control, commonly adopted storage methods do not ensure long-term preservation of sensory and functional quality.
A limitation of the present study lies in the use of conventional microbiological analyses, which provide quantitative information but limited insight into microbial community dynamics.
Future research should integrate these methods with advanced microbial profiling approaches, such as metagenomic or metataxonomic sequencing, to better elucidate microbial shifts associated with spoilage processes and sensory degradation.
Acknowledgments
The authors are grateful to Gilberto Bragato of the CREA-Viticulture and Enology of Gorizia (Italy), who contributed to elaborating Figure 1, to Martina Muzzalupo, who contributed to elaborating the Graphical Abstract, and to the other people who contributed to the fungi material collection. Particular thanks are also due to the judges of CibusLAB - Sensory analysis laboratory for the protection and enhancement of Italian agri-foods, Cosenza, Italy, for having conducted the sensory analysis on the truffles.
Consent for Publication
All authors have read and agreed to the published version of the manuscript.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Mandatory Disclosure on Use of Artificial Intelligence
The authors declare that AI-assisted tools were used for linguistic revision and stylistic editing of the manuscript, specifically OpenAI ChatGPT (GPT-5.5). All scientific content and references were manually verified by the authors for accuracy, relevance, and consistency with the final version of the manuscript.
Author Contribution
AC: Funding acquisition, Conceptualization, Supervision, Validation, Writing – review & editing. VS: Methodology, Data curation. AG: Methodology, Formal analysis. RT: Validation, Methodology, Formal analysis. RP: Formal analysis, Data curation. MP: Methodology, Formal analysis. KG: Formal analysis. IM: Funding acquisition, Project administration, Supervision, Writing – review & editing. MRL: Project administration, Supervision, Writing – review & editing. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interests
The authors declare no conflicts interests.
Funding
This work was supported by an ex-60% grant from the University of Calabria; MICROLIVE Project –PSR Calabria 2014–2020, Measure 16, Intervention 16.02.01: “Support for pilot projects and the development of new products, practices, processes and technologies in the agri-food and forestry sector (CUP: J96G20000340005); VIVATUCA Project, “Development of Nurseries for Truffle-Mycorrhized Forest Plants in Calabria: An Opportunity for the Ecological and Economic Enhancement of the Territory” (CUP: C23C25001170002).
REFERENCES
Al-Laith, A., 2010. Antioxidant components and antioxidant/-antiradical activities of desert truffle (Tirmania nivea) from various Middle Eastern origins. Journal of Food Composition and Analysis. 23: 15–22. 10.1016/J.JFCA.2009.07.005
Alothman, M., Bhat, R., and Karim, A.A., 2009. Effects of radiation processing on phytochemicals and antioxidants in plant-produce. Trends in Food Science and Technology. 20: 201–212. 10.1016/j.tifs.2009.02.003
Altamim, E.A., 2024. Gamma irradiation-mediated alterations in chemical composition and antimicrobial efficacy of Zubaidi, a wild edible desert truffle (Terfezia boudieri), with implications for pharmaceutical applications. Food Science and Technology (Campinas). 44: e00251. 10.5327/fsct.00251
Amadoro, C., Tremonte, P., Succi, M., Tipaldi, L., Pannella, G., Coppola, R., et al. 2013. Chitosano, ultrasuoni e atmosfera-modificata per la conservazione del tartufo nero (Tuber aestivum Vitt.) fresco. Industrie Alimentari. 52: 25–32.
Amicucci, A., Zambonelli, A., Giomaro, G., Potenza, L., and Stocchi, V., 1998. Identification of ectomycorrhizal fungi of the genus Tuber by species-specific ITS primers. Molecular Ecology. 7: 273–277. 10.1046/j.1365-294X.1998.00357.x
Baral, H.O., Weber, E., Gams, W., Hagedorn, G., Liu, B., Liu, X., et al. 2018. Generic names in the Orbiliaceae (Orbiliomycetes) and recommendations on which names should be protected or suppressed. Mycological Progress. 17: 5–31. 10.1007/s11557-017-1300-6
Bayram, I., and Decker, E.A., 2023. Underlying mechanisms of-synergistic antioxidant interactions during lipid oxidation. Trends in Food Science and Technology. 133: 219–230. 10.1016/j.tifs.2023.02.003
Benaceur, F., Chaibi, R., Berrabah, F., Neifar, A., Leboukh, M., Benaceur, K., et al. 2020. Purification and characterization of latent polyphenol oxidase from truffles (Terfezia arenaria). International Journal of Biological Macromolecules. 145: 885–893. 10.1016/j.ijbiomac.2019.09.126
Bonito, G., 2009. Fast DNA-based identification of the black truffle Tuber melanosporum with direct PCR and species-specific primers. FEMS Microbiology Letters. 301: 171–175. 10.1111/j.1574-6968.2009.01812.x
Bruno, L., Chiappetta, A., Muzzalupo, I., Gagliardi, C., Iaria, D., Bruno, A., et al. 2009. Role of geranylgeranyl reductase gene in organ development and stress response in olive (Olea europaea) plants. Functional Plant Biology. 36: 370–381. 10.1071/FP08219
Çayan, F., Tel-Çayan, G., Deveci, E., Duru, M.E., and Öztürk, M., 2022. Isolation and identification of compounds from truffle Reddellomyces westraliensis and their antioxidant, cytotoxic and enzyme inhibitory activities. Process Biochemistry. 121: 553–562. 10.1016/j.procbio.2022.08.001
Chiappetta, A., Muto, A., Muzzalupo, R., and Muzzalupo, I., 2017. New rapid procedure for genetic characterization of Italian wild olive (Olea europaea) and traceability of virgin olive oils by means of SSR markers. Scientia Horticulturae. 226: 42–49. 10.1016/j.scienta.2017.08.022
Culleré, L., Ferreira, V., Venturini, M.E., Marco, P., and Blanco, D., 2012. Evaluation of gamma and electron-beam irradiation on the aromatic profile of black truffle (Tuber melanosporum) and summer truffle (Tuber aestivum). Innovative Food Science and Emerging Technologies. 13: 151–157. 10.1016/j.ifset.2011.09.003
Culleré, L., Ferreira, V., Venturini, M.E., Marco, P., and Blanco, D., 2013. Chemical and sensory effects of the freezing process on the aroma profile of black truffles (Tuber melanosporum). Food Chemistry. 136: 518–525. 10.1016/j.foodchem.2012.08.030
Daei, B., Azadmard-Damirchi, S., Javadi, A., and Torbati, M., 2023. Effects of mild thermal processing and storage conditions on the quality attributes and shelf life of truffles (Terfezia claveryi). Foods. 12: 2212. 10.3390/foods12112212
Epping, R., Lisec, J., and Koch, M., 2024. Changes in Black Truffle (Tuber melanosporum) aroma during storage under different conditions. Journal of Fungi. 10: 354. 10.3390/jof10050354
Gan, Z., Yang, J., Tao, N., Yu, Z., and Zhang, K.Q., 2007. Cloning and expression analysis of a chitinase gene crchi1 from the mycoparasitic fungus Clonostachys rosea (syn. Gliocladium roseum). Journal of Microbiology. 45: 422–430.
Gryndler, M., Hršelová, H., Soukupová, L., Streiblová, E., Valda, S., Borovička, J., et al. 2011. Detection of summer truffle (Tuber aestivum Vittad.) in ectomycorrhizae and in soil using specific primers. FEMS Microbiology Letters. 318 : 84–91. 10.1111/j.1574-6968.2011.02243.x
Guo, T., Wei, L., Sun, J., Hou, C., and Fan, L., 2011. Antioxidant activities of extract and fractions from Tuber indicum Cooke & Massee. Food Chemistry. 127: 1634–1640. 10.1016/J.FOODCHEM.2011.02.030
Hajjar, S.E., Massantini, R., Botondi, R., Kefalas, P., and Mencarelli, F., 2010. Influence of high carbon dioxide and low oxygen on the postharvest physiology of fresh truffles. Postharvest Biology and Technology. 58: 36–41. 10.1016/j.postharvbio.2010.04.008
Hall, I., Brown, G., and Zambonelli, A., 2007. Taming the truffle. The history, lore, and science of the ultimate mushroom. Portland, OR, USA: Timber Press. Available from: https://books.google.it/books/about/Taming_the_Truffle.html?id=taQGqqcaEYoC&redir_esc=y
Healy, R., Bonito, G.M., and Smith, M.E., 2016. A brief overview of the systematics, taxonomy, and ecology of the Tuber rufum clade. In Zambonelli, A., Iotti, M., Murat, C. (Eds.). True truffle (Tuber spp.) in the world. New York: Springer. pp. 125–136. https://link.springer.com/chapter/10.1007/978-3-319-31436-5_8
Henrion, B., Chevalier, G., and Martin, F., 1994. Typing truffle species by PCR amplification of the ribosomal DNA spacers. Mycology Research. 98: 37–43. 10.1016/S0953-7562(09)80333-X
Horwitz, W., 2000. Official methods of analysis of AOAC International. Gaithersburg, Md: Ed. Association of Official Analytical Chemists. Available from: https://search.worldcat.org/it/title/Official-methods-of-analysis-of-AOAC-International/oclc/796380460
Iotti, M., Amicucci, A., Bonito, G., Bonuso, E., Stocchi, V., and Zambonelli, A., 2007. Selection of a set of specific primers for the identification of Tuber rufum: A truffle species with high genetic variability. FEMS Microbiology Letters. 277: 223–231. 10.1111/j.1574-6968.2007.00963.x
Lanfranco, L., Wyss, P., Marzachi, C., and Bonfante, P., 1993. DNA probes for identification of the ectomycorrhizal fungus Tuber magnatum Pico. FEMS Microbiology Letters. 114: 245–251. 10.1111/j.1574-6968.1993.tb06581.x
Lee, H., Nam, K., Zahra, Z., and Farooqi, M.Q.U., 2020. Potentials of truffles in nutritional and medicinal applications: A review. Fungal Biology and Biotechnology. 7: 1–17. 10.1186/s40694-020-00097-x
Leonardi, P., Murat, C., Puliga, F., Iotti, M., and Zambonelli, A., 2020. Ascoma genotyping and mating type analyses of mycorrhizas and soil mycelia of Tuber borchii in a truffle orchard established by mycelial inoculated plants. Environmental Microbiology. 22: 964–975. 10.1111/1462-2920.14777
Li, Q., Hu, H., Tan, X., Wang, J., Mei, R., Jiang, F., et al. 2024. Effects of storage in an active and spontaneous controlled O2/CO2 atmosphere on volatile flavor components and the microbiome of truffles. ACS Omega. 9: 9331–9347. 10.1021/acsomega.3c08375
Liu, T., Feng, T., and Chen, W., 2017. Identification of volatile flavour components of Tuber melanosporum using simultaneous distillation-extraction. Czech Journal of Food Sciences. 35: 483–487. 10.17221/31/2017-CJFS
Loizzo, M.R., Tundis, R., Leporini, M., D’Urso, G., Gagliano Candela, R., Falco, T., et al. 2021. Almond (Prunus dulcis cv. Casteltermini) skin confectionery by-products: New opportunity for the development of a functional blackberry (Rubus ulmifolius Schott) jam. Antioxidants. 10: 1218. 10.3390/antiox10081218
Mannozzi Torini, L., 1994. Il tartufo e la sua coltivazione. Edagricole, Bologna, Italy. pp. 1–106 Available from: https://opac.sbcividalese.it/ricerca/dettaglio/il-tartufo-e-la-sua-coltivazione/40289
Massantini, R., Moscetti, R., Contini, M., and Bellincontro, A., 2009. Confezionamento in film plastico di zucchino (Cucurbita pepo L.) parzialmente processato. Industrie Alimentari. 48: 26–37.
Mello, A., Murat, C., and Bonfante, P., 2006. Truffles: Much more than a prized and local fungal delicacy. FEMS Microbiology Letters. 260: 1–8. 10.1111/j.1574-6968.2006.00252.x
Paolocci, F., Rubini, A., Riccioni, C., Topini, F., and Arcioni, S., 2004. Tuber aestivum and Tuber uncinatum: Two morphotypes or two species? FEMS Microbiology Letters. 235: 109–115. 10.1111/j.1574-6968.2004.tb09574.x
Patel, S., 2012. Food, health and agricultural importance of truffles: A review of current scientific literature. Current Trends in Biotechnology and Pharmacy. 6: 15–27.
Pegler, D.N., Spooner, B.M., and Young, T.W.K., 1993. British truffles. A revision of british hypogeous fungi. London, United Kingdom: Royal Botanic Garden, Kew Publishing Royal Botanic Gardens Publication. pp. 226.
Phong, W.N., Chang, S., Payne, A.D., Dykes, G.A,., and Coorey, R., 2022. Microbiological evaluation of whole, sliced, and freeze-dried black truffles (Tuber melanosporum) under vacuum packaging and refrigerator storage. JSFA Reports. 2: 92–99. 10.1002/jsf2.32
Reale, A., Sorrentino, E., Iacumin, L., Tremonte, P., Manzano, M., Maiuro, L., et al. 2009. Irradiation treatments to improve the shelf life of fresh black truffles (truffles preservation by gamma-rays). Journal of Food Science. 74: M196–200. 10.1111/j.1750-3841.2009.01142.x
Rivera, C.S., Blanco, D., Oria, R., and Venturini, M.E., 2010. Diversity of culturable microorganisms and occurrence of Listeria monocytogenes and Salmonella spp. in Tuber aestivum and Tuber melanosporum ascocarps. Food Microbiology. 27: 286–293. 10.1016/j.fm.2009.11.001
Rivera, C.S., Blanco, D., Marco, P., Oria, R., and Venturini, M.E., 2011. Effects of electronbeam irradiation on the shelf life, microbial populations and sensory characteristics of summer truffles (Tuber aestivum) packaged under modified atmospheres. Food Microbiology. 28: 141–148. 10.1016/J.FM.2010.09.008
Rivera, C.S., Venturini, M.E., Marco, P., Oria, R., and Blanco, D., 2011b. Effects of electronbeam and gamma irradiation treatments on the microbial populations, respiratory activity and sensory characteristics of Tuber melanosporum truffles packaged under modified atmospheres. Food Microbiology. 28: 1252–1260. 10.1016/j.fm.2011.05.002
Rosnes, J.T., Sivertsvik, M., and Skara, T., 2003. Combining MAP with other preservation techniques. In Ahvenainen, R. (Ed.). Novel food packaging techniques. Cambridge, UK: Woodhead Publishing. pp. 287–304.
Saltarelli, R., Ceccaroli, P., Cesari, P., Barbieri, E., and Stocchi, V., 2008. Effect of storage on biochemical and microbiological parameters of edible truffle species. Food Chemistry. 109: 8–16. 10.1016/j.foodchem.2007.11.075
Savini, S., Loizzo, M.R., Tundis, R., Mozzon, M., Foligni, R., Longo, E., et al. 2017. Fresh refrigerated Tuber melanosporum truffle: Effect of the storage conditions on the antioxidant profile, antioxidant activity and volatile profile. European Food Research and Technology. 243: 2255–2263. 10.1007/s00217-017-2927-x
Savini, S., Longo, E., Servili, A., Murolo, S., Mozzon, M., Romanazzi, G., et al. 2020. Hypobaric packaging prolongs the shelf life of refrigerated black truffles (Tuber melanosporum). Molecules. 25: 3837. 10.3390/molecules25173837
Sbrana, C., Agnolucci, M., Bedini, S., Lepera, A., Toffanin, A., Giovannetti, M., et al. 2002. Diversity of culturable bacterial populations associated to Tuber borchii ectomycorrhizas and their activity on T. borchii mycelial growth. FEMS Microbiology Letters. 211: 195–201. 10.1016/S0378-1097(02)00712-7
Shen, Q., Kong, F., and Wang, Q., 2006. Effect of modified atmosphere packing on the browning and lignification of bamboo shoots. Journal of Food Engineering. 77: 348–354. 10.1016/j.jfoodeng.2005.06.041
Sicari, V., Tundis, R., Romeo, R., Reitano, A., Belsito, E.L., Leggio, A., et al. 2024. Evaluation of the shelf life of ready-to-eat fresh Bamboo sprouts (Phyllostachys edulis) packaged in a-modified atmosphere or vacuum: A comparative study. Antioxidants. 13: 185. 10.3390/antiox13020185
Spanò, F.M., and Mattia Serafin, C., 2024. Il tartufo nero di Calabria: L’inizio di un nuovo racconto. Roma, Italy: Gangemi Ed. pp. 1–256.
Strojnik, L., Grebenc, T., and Ogrinc, N., 2020. Species and geographic variability in truffle aromas. Food and Chemical Toxicology. 142: 111434. 10.1016/j.fct.2020.111434
Tejedor-Calvo, E., Morales, D., García-Barreda, S., Sánchez, S., Venturini, M.E., Blanco, D., et al. 2020. Effects of gamma irradiation on the shelf-life and bioactive compounds of Tuber aestivum truffles packaged in passive modified atmosphere. International Journal of Food Microbiology. 332: 108774. 10.1016/j.ijfoodmicro.2020.108774
Tejedor-Calvo, E., Amara, K., Reis, F.S., Barros, L., Martins, A., Calhelha, R.C., et al. 2021. Chemical composition and evaluation of antioxidant, antimicrobial and antiproliferative activities of Tuber and Terfezia truffles. Food Research International. 140: 110071. 10.1016/j.foodres.2020.110071
Tejedor-Calvo, E., Baquero-Aznar, V., Vega-Diez, S., Salvador, M., Sanz, M., Sánchez, S., et al. 2024. Potential of a smart gelatine hydrogel-based package to extend fresh black truffle (Tuber melanosporum) shelf-life preserving its aroma profile. Food Hydrocolloids. 151: 109874. 10.1016/j.foodhyd.2024.109874
Tenuta, M.C., Deguin, B., Loizzo, M.R., Dugay, A., Acquaviva, R., Malfa, G.A., et al. 2020. Contribution of flavonoids and iridoids to the hypoglycaemic, antioxidant, and nitric oxide (NO) inhibitory activities of Arbutus unedo L. Antioxidants. 9: 184. 10.3390/ANTIOX9020184
White, T.J., Bruns, T., Lee, S., and Taylor, J., 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J. (Eds.). PCR protocols: A guide to methods and applications. New York, USA: Academic Press. pp. 315–322.
Yan, X., Wang, Y., Sang, X., and Fan, L., 2017. Nutritional value, chemical composition and antioxidant activity of three Tuber species from China. AMB Express. 7: 136. 10.1186/s13568-017-0431-0
Zambonelli, A., Iotti, M., Murat, C., eds. 2016. True truffle (Tuber spp.) in the world: Soil ecology, systematics and biochemistry. Springer, Cham. https://link.springer.com/book/10.1007/978-3-319-31436-5
Supplementary
Table S1. Evolution of pH in truffles stored in two different conditions.
| Truffles | pH | Sign | ||||
|---|---|---|---|---|---|---|
| t0 | t7 | t14 | t21 | t28 | ||
| TUUR | 6.67a | 6.46b | 6.34bc | 6.14cd | 5.86d | ** |
| TUUV | 6.67a | 6.40b | 6.37b | 6.17c | 6.01c | ** |
| TMUR | 6.64a | 6.35b | 6.55a | 6.20b | 6.03c | ** |
| TMUV | 6.64a | 6.56a | 6.34b | 6.24b | 5.86c | ** |
| TBUR | 6.69a | 6.54ab | 6.37b | 6.32b | 6.23c | ** |
| TBUV | 6.69a | 6.56ab | 6.52ab | 6.36b | 6.12c | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TMUR: Tuber mesentericum under rice; TBUR: Tuber borchii under rice, and TUUV: T. aestivum var. uncinatum under vacuum; TMUV: T. mesentericum under vacuum; TBUV: T. borchii under vacuum. Data are expressed as means ± SD (n = 3). Differences between samples were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant. **P ≤ 0.01.
Table S2. Total microbial count (TBC) in truffles stored in two different conditions.
| CFU/g | t0 | t7 | t14 | t21 | t28 | Sign. |
|---|---|---|---|---|---|---|
| TUUR | 1.18 × 106a | 2.79 × 107b | 3.5 × 1010e | 2 × 109d | 1.9 × 107c | ** |
| TUUV | 1.3 × 106a | 1.89 × 107b | 4.1 × 109e | 1.92 × 108c | 1.4 × 109d | ** |
| TMUR | 7.8 × 106b | 3.07 × 107c | 3.2 × 109e | 2.4 × 106a | 1.7 × 107d | ** |
| TMUV | 6.0 × 106a | 9.7 × 106b | 3.3 × 109e | 2.49 × 109d | 1.53 × 109c | ** |
| TBUR | 1.4 × 106a | 4.42 × 107b | 2.9 × 109d | 4.3 × 109e | 2.7 ×109c | ** |
| TBUV | 1 × 106a | 1.51 × 107b | 2.5 × 109d | 2.9 × 109e | 1.2 × 108c | ** |
TUUR: Tuber aestivum var. uncinatum under rice; TMUR: Tuber mesentericum under rice; TBUR: Tuber borchii under rice, and TUUV: T. aestivum var. uncinatum under vacuum; TMUV: T. mesentericum under vacuum; TBUV: T. borchii under vacuum. Data are expressed as means ± SD (n = 3). Differences between samples were evaluated by one-way ANOVA followed by Tukey’s post hoc test. Results followed by letters are significant. **P ≤ 0.01.
Table S3. Degradation Kinetics of TPC and TFC during truffle’s storage.
| Bioactives | Treatment | Reaction order | K-value | Equation | R2 | Half-life (t1/2) (Days) |
|---|---|---|---|---|---|---|
| TPC | ||||||
| TUUR | 0 | 0.4614 | y = −0.4614x + 15.34 R2 = 0.973 | 0.973 | 1.5020 | |
| TUUV | 0 | 0.3243 | y = −0.3243x + 13.62 R2 = 0.7255 | 0.7255 | 2.1369 | |
| TMUR | 0 | 0.3386 | y = −0.3386x + 16.23 R2 = 0.9746 | 0.9746 | 2.0467 | |
| TMUV | 0 | 0.3557 | y = −0.3557x + 14.74 R2 = 0.9602 | 0.9602 | 1.9483 | |
| TBUR | 0 | 0.35 | y = −0.35x + 16.02 R2 = 0.9683 | 0.9683 | 1.9800 | |
| TBUV | 0 | 0.35 | y = −0.35x + 15.97 R2 = 0.9857 | 0.9857 | 1.9800 | |
| TFC | ||||||
| TUUR | 0 | 0.1223 | y = −0.1223x + 5.918 R2 = 0.8864 | 0.8864 | 5.6664 | |
| TUUV | 0 | 0.1234 | y = −0.1234x + 5.972 R2 = 0.8021 | 0.8021 | 5.6158 | |
| TMUR | 0 | 0.1626 | y = −0.1626x + 5.984 R2 = 0.9825 | 0.9825 | 4.2620 | |
| TMUV | 0 | 0.088 | y = −0.088x + 5.798 R2 = 0.8709 | 0.8709 | 7.875 | |
| TBUR | 0 | 0.0777 | y = −0.0777x + 5.152 R2 = 0.9951 | 0.9951 | 8.9189 | |
| TBUV | 0 | 0.1283 | y = −0.1283x + 6.184 R2 = 0.9128 | 0.9128 | 5.4014 |
TUUR: Tuber aestivum var. uncinatum under rice; TMUR: Tuber mesentericum under rice; TBUR: Tuber borchii under rice, and TUUV: T. aestivum var. uncinatum under vacuum; TMUV: T. mesentericum under vacuum; TBUV: T. borchii under vacuum.